Цитология методичка 1-15 сабак. Саба 1 Апаратты дидактикалы топтама
 Скачать 1.99 Mb. Скачать 1.99 Mb.
|

БелоктардыңпосттрансляциялықмодификацияларыКейбір белоктар синтез аяқталғаннан кейін құрылымдық өзгерістерге ұшырайды. Әдетте бірқатар белоктарда ыдырау немесе тримминг байқалады. Мысалы, инсулиннің (қызмет атқаруы) активтілігі геннің бірінші реттік өнімі ретінде синтезделген проинсулиннің орталық бөлігін ферменттер көмегімен алып тастау арқылы қамтамасыз етіледі. Көпшілік белоктардың активтілігі олардың басқа, құрылысы ұқсас немесе ұқсамайтын әртүрлі белоктармен байланысқан жағдайда іске асырылады. Мысалы, қызмет атқаруға қабілетті гемоглобин гемнен және глобиндік тізбектерден тұрады. Бұл байланыс бұзылса ағза темір жетіспейтін анемия ауруына шалдығады. ГендерактивтілігініңгормональдықреттелуіЖоғары сатыдағы өсімдіктер мен жануарларда жасушааралық өзара әрекеттесулер олардың тіршілік үдерісінде маңызды роль атқарады. Жасушалардың өзара әрекеттесуі гормональдық әсер етуге байланысты іске асырылады. Гормондар тікелей жасуша – нысанаға немесе ұлпа – нысанаға әсер етеді. Пептидтік гормондар (инсулин эпинефрин т.б.) өздерінің ірі молекулалы болуына байланысты жасуша ішіне ене алмайды. Олар жасушаға рецепторлық белоктармен әрекеттесу арқылы әсер етеді. Рецепторлық белоктар әдетте нысаналық жасушаның мембраналарында орналасады. Осылай әсер ету арқылы пептидтік гормондар жасушаішілік циклдың АМФ (цАМФ)-тың деңгейін реттейді. Циклдық АМФ протеин-киназа ферментін активтендіреді. Ал протеин-киназа ферменті көптеген арнайы қызмет атқаратын ферменттерді фосфорилдейді (активтендіріледі). Стероидтық гормондар (эстроген, прогестерон, тестерон, экдизон) жасуша ішіне оңай еніп, жасуша-нысана цитоплазмасындағы арнайы рецепторлық белоктармен берік байланысады. Түзілген гормон + рецепторлық белок кешені арнайы гендердің транскрипциясын активтендіреді. Оның екі жолы бар: 1. Гормон + рецепторлық белок кешені арнайы гистонды емес белоктармен өзара әрекеттесіп нақты гендердің транскрипциясын активтендіреді. 2. Гормон + рецепторлық белок кешені гендердің энхансерлік және промоторлық учаскелерінде орналасқан арнайы нуклеотидтер жүйесімен байланысып сол гендердің транскрипциясын активтендіреді (R.M. Evans, 1988). Осы екі жағдайда да гормон + протеиндік кешен транскрипция үдерісіне прокариоттардағы САР-с АМР-кешенінің әсеріне ұқсас, позитивтік реттеуші әсер көрсетеді. Олай болса, гендер активтілігі жасушалардың (ағзалардың) нақты тіршілік жағдайларына байланысты болып, тұқым қуалау ақпаратының жүзеге асырылуының барлық кезеңдерінде түрлі механизмдермен бақыланады. Сабақ 12 Жасушалық цикл кезеңдерінің ретімен алмасып отыруы циклин және циклин-тәуелді киназа (Ц-ЦТК) кешендері, белок-ферменттерінің әсері арқылы іске асырылады. Циклиннің ЦТК-ға жалғануы ЦТК-ны ғана активтендіріп қоймай, басқа да белоктарға әсері етеді. Жасшадағы циклиннің концентрациясы жасушалық цикл барысында циклды түрде өзгеріп отырады. Жасушалық цикл ЦТК (Cdk) активтілігінің өзгеріуімен оның реттеуші суббірлігі болып табылатын циклиннің циклға сай түзіліп және қайта ыдырап отыруымен бақыланады. Түрлі циклиндердің синтезделіп және қайта ыдырап отыруы жасушалық циклдың фазаларының ретімен алмасып отыруын қамтамасыз етеді. Жасушалық циклдың түрлі фазаларында әртүрлі циклиндер түзіледі де ЦТК-мен жалғанып Ц-ЦТК кешендер түзеді. Бұл кешенде циклин активтендіруші , ал ЦТК -лар катализаторлар қызметін атқарады (сурет 1). Олай болса төменде көрсетілген үдерістер жүргенде ғана митоздық циклдың реттелуінің бақылануы іске асырылады: 1. циклинге тәуелді киназа (Cdk) тек өзіне тән циклинмен қосылғанда ғана әсерін көрсетеді 2. Cdk фосфорилденгенде және дефосфорилденгенде 3.INK4 және KIP1 тұқымдасындағы арнайы ингибиторлық белоктармен әсерлескенде. Сонымен Ц-ЦТК кешендерінің механизмдері жалпы белгілі бір ретпен жүретін төмендегі үдерістерді реттейді: а) митоздық циклдың алдыңғы сатысын реттеуші факторлардың активсізденуін; б) митоздық циклдың осы кезеңін қамтамасыз ететін факторлардың активтенуін; в) жасуша бөлінуінің келесі кезеңіне қажетті факторлар кешенінің түзілуімен активтенуін. 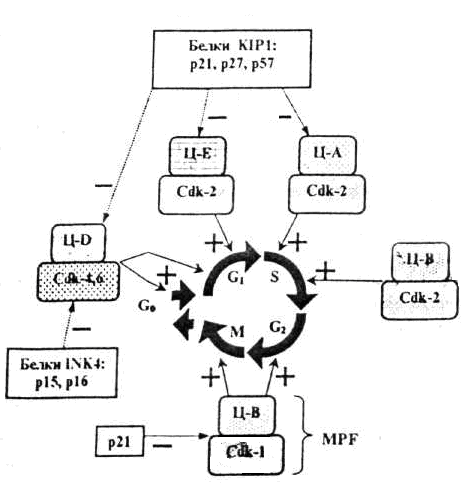 С. 1. Жасушалық циклдың түрлі фазаларын анықтайтын циклин- Cdk кешені Ц- циклиндер, Cdk – циклин-тәуелді киназалар, MPF –митозстимулдаушы фактор. Төртбұрыш қоршауларда – ингибиторлар кешені. Жасушаның бөлінуінің басталуы үшін сырқы орта жағдайдың қолайлы болуымен қатар арнайы сигналдық молекулалар – өсу факторлары (митогендер) және жасушаның қандайда бір беткейге бекінуі (эпителиоциттердің базальдық мембранаға, фибробластардың коллаген талшықтарына жалғануы) қажет және жасуша беткейіндегі рецепторлармен байланысқанда және контакталық тежелу болмаған жағдайда митоздық цикл механизмі іске қосылады. Жасушалық циклді реттеудің сигналдық жолдары Митотикалық цикл үдерісінің іске қосылуы циклин Д-ЦТК4 және/немесе циклин Д-ЦТК-6 кешенінің әсеріне байланысты болады. Бастаушы кешенді белсендіру үшін жасушадан тыс реттегіштер – өсу факторынан, яғни өсудің эпидермалдық факторынан (ЭФР) және антиген саналатын жасушадан (АПК) сигналдардың түсуі қажет. Тирозинкиназа рецепторымен қабылданатын сигналдар каскадтық жолмен киназалық домен рецепторларын, митогенді активтендіреті протеинкиназаларды (МАПК) белсендіреді, олар митогеннің әсеріне тез әрі ерте жауап беретін (FOS, JUN) гендері үшін транскрипциялық факторлар (ТФ) болып табылады. Ерте жауап беретін (FOS, JUN) гендерінің өнімдері өз кезегінде баяу жауап гендері үшін ТФ болады және оларды белсендіреді. Баяу жауап беретін ген өнімдері, оның ішінде ақуыз-ферменттер, циклин Д-ЦТК 4, ЦТК 6, митоздық циклдың G1–кезеңінің бірінші жартысында әсер ететін циклин-ЦТК кешендерінің пайда болуына әкеледі. Сонымен қатар басқа да гендер, оның ішінде ақуыз Мус геніде белсенді болады. Мус ақуызы р27 генінің белсенділігін тежейді, р27 ақуызы циклин-ЦСК кешендерін тежейді және Сdc25a генін белсендіреді, оның өнімі спецификалық фосфатаза киназа мен ЦЗК2 фосфорилдеу арқылы оларды белсендіреді. Бұл үдерістердің нәтижесі әкеледі: 1) жасушадағы циклин-Д және ЦТК4, ЦТК 6 киназаларының көбеюіне; 2) киназа (Cdk) ингибиторларының азюына; Осының барлығы жасушада белсенді циклин-Д-ЦТК4,6-ы кешендерінің жеткілікті мөлшерін қамтамасыз етеді. Осылайша, G1 кешендері: а) G1-фазада орын алатын үдерістерді; б) ДНҚ репликациясы үдерісіне қатысатын факторлар мен ферменттердің синтезін реттейді. 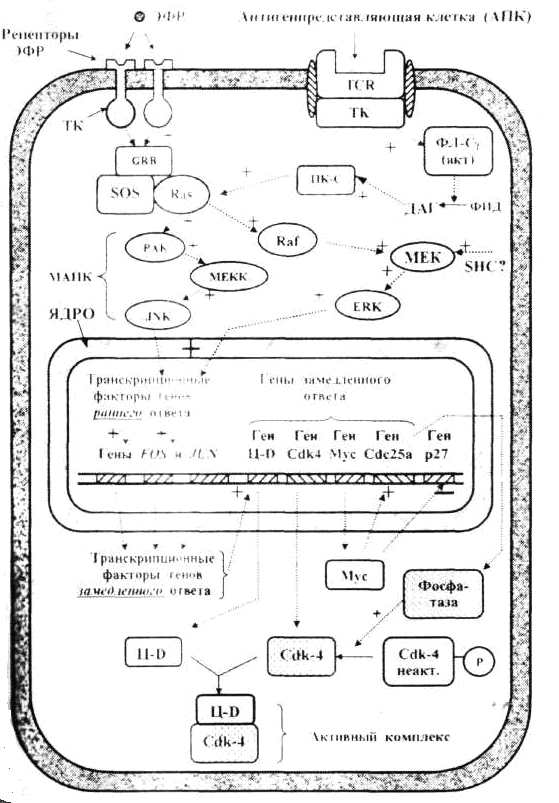 2-сурет. Жасуша пролиферациясын инициялайтын сигналдық жолдар: а) бағаналық эрпителиоциттер (басталуы -ЭФР); б) Т-мимфоциттер (АПК-мен өзара әрекеттесуден басталады); S-және G2-кезеңдерінің кешендерінің әсері S-кезеңнің кешені ЦА-Cdk2 және ЦВ-Cdk2 болып табылады. Бұл кешендердің негізгі міндеті-ДНҚ-ның репликациясын яғни әрбір учаскесінің тек бір рет ғана репликациядан өтуін қамтамасыз етеді. Цитоплазмада S-кезеңінде центриольдердің екі еселенуі де жүреді. G2-кезеңде MPF (митозды стимулдаушы фактор –mitosis-promoting factor) жинақталуы орын алады. MPF- циклин В-Cdk1.Cdk1кешені.G2-кезеңінде бөлінуі ұршығын түзетін тубулин ақуызы синтезделеді. Митоздың реттелуі Профаза және метафаза сатыларында MPF факторының белсенділігі жоғары болады, маңызды роль атқарады. Анафазаның өту үшін MPF бұзылуы қажет. Бұл анафазаны қамтамасыз ететін фактормен жүзеге асырылады (АРС –G1 кезеңінде синтезделген). Митоздың метафазасында MPF басқа ақуыздармен қатар APC –ті де фосфорлайды және, ол іске қосылады да MPF құрамына кіретін циклин-В молекулаларын бұза бастайды, және MPF профазамен метафазада фосфорланған ақуыздар қайта дефосфорланады. Бұл ақуыздардың әсері жасушаның құрылымдарды қалпына келтіреді және цитотомияны іске қосады (цитокинезді). Митозды стимулдаушы факторлармен қатар, жасушаның митозға енуіне кедергі келтіретін қарсы факторларда бар: 1) ісіктер некрозының факторы (ІНФ) – МАПК каскадының тежелуіне ықпал етеді,осының салдарынан Д - ЦЗК4, 6 циклиннің белсенді кешендерінің саны төмендейді. 2) өсудің трасформациалаушы факторы (ӨТФ) - smad тұқымдасының цитоплазмалық ақуыздарының каскадтық механизмдерін іске қосады. Бұл кешен р15 және р21 ақуыздарының синтезін кодтайтын гендер үшін транскрипциялық фактор рөлін атқарады яғни ЦТК-киназалардың ингибиторлары болып табылады. Осы ингибиторлардың жасушада жиналуы пролиферацияның тежелуіне әкеледі. 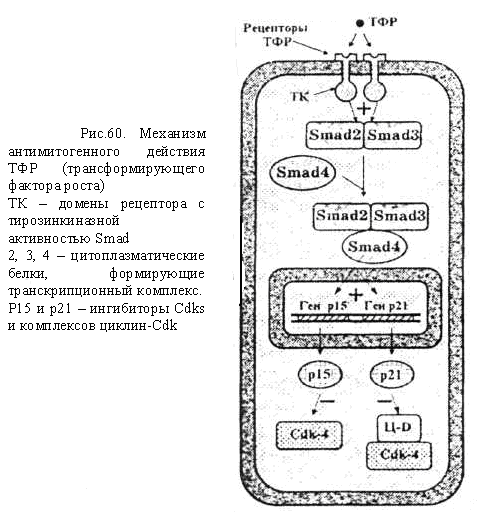 ДНҚ зақымдануын бақылайтын чекпойнттар Митотикалық циклді бақылайтын және реттейтін үдерістермен қатар жасушада өз жағдайын өзі бақылау үдерісіде жүреді. Генетикалық материалдың репликациясы мен таралуының дұрыстығын бақылау митотикалық циклдің (G1, S, G2 және М-фазалар) төрт кезеңінде жүзеге асырылады және арнайы жасушалық механизмдермен (р53 ақуызымен) бақыланады. Кесте Тексеру жүретін пункттерді салыстырып тексеру нүктелері (check-points):
Егер осы фазалардың кез келгенінде зақымданулар байқалса, жасушадағы репарация ферменттері зақымдалған ДНҚ бөліктерін қалпына келтірілгенге дейін жасуша тиісті фазаларда кідіреді. Фазадағы кідіріс зақымдалған негіздердің көшірілуін болдырмайды және ДНҚ мутациясын тежейді. Циклді тоқтатуда р53 ақуызының маңызы зор. Ол жасушада үнемі синтезделіп отырады, бірақ өте тез жойылады. Жасушадағы ақауларды жою кезінде р53 ақуыз концентрациясы төмендейді, Cdk-ның ингибиторы ажырайды да Cdk қайтадан экспрессияланады. Осылайша, "тексеру" нәтижелеріне байланысты үш нұсқаның бірі таңдалады, бақылыу былай аяқталуы мүмкін: а) зақымданулар болмаған кезде-циклдің келесі кезеңіне тоқтаусыз өтеді; б) зақымданулар анықталған жағдайда ақауларды түзету үшін жасуша ағымдағы кезеңде кідіреді; в) егер анықталған бұзылулар ығыстырлмаса апоптоз механизмі іске қосылады. 12-сабақ: Өзгергіштік. Геномдық және хромосомалық мутациялар Өзгергіштік – бұл орта факторларының әсерінен тірі ағзалардың өзгеруі және жаңа қасиеттер ие бола алу қабілеті. Фенотиптік (тұқымқуаламайтын) және генотиптік (тұқымқуалайтын) өзергіштік деп ажыратылады. Тұқым қуаламайтын немесе модификациялық өзгергіштік ағзалардың түрлі қоршаған орта факторларынан (температур, ылғал және т.с.с.) ағзалардың өзгере алу қабілеті. Бұл өзгергіштік типі генотипттің өзгеруіне байланысты емес, сондықтан тұқым қуаламайды. Бірақ, кез-келген белгінің модификациялық өзгеру шегі, яғни норма реакциясы генотип арқылы беріледі. Сонымен қатар, тұқым қуаламайтын өзгергіштікке фенокопиялар мен морфоздар жатады. Фенокопиялар — сыртқы ортаның қолайсыз факторларының әсерінен мутацияға ұқсас фенотиптегі өзгерістер. Фенокопиялардың себептері: ұрықта оттегі жеткізіксіздігі (орталық жүйке жүйесінің зақымдалуы); инфекциялық аурулар (токсоплазмоз, қызылша, сифилис); эндокриндік бұзылыстар; психикалық травма және эмоционалдық күйзеліс; цитотоксикалық немесе антиметоболиттік әсерлері бар дәрілік заттар (талидомид); микроэлементтердің (Fe, Co, Cu), дәрумендердің (С, Е, В1, РР және т.б.) жетіспеуі; алкоголизм; жүктілікке қарсы (контрацептивтерді) және жүктілікті үзетін дәрілік заттарды сауатсыз қолдану. Медицинада фенокопиялар — бұл тұқым қуаламайтын аурулар. Морфоздар – экстремалды немесе ерекше болып табылатын орта факторлары әсерінен болған тұқымқуаламайтын өзгерістер. Морфоздар сәулелену әсерінен болса - рентгеноморфоздар, ал химиялық заттар әсерінен болса—хемоморфоздар деп аталады. Морфоздарға тән ерекшеліктер: тұқымқуаламайды, адаптивті емес, әдетте, қайта қалпына келмейді. Мысалы: күйіктер, жарақаттар және т.б. Генотиптік немесе тұқымқуалайтын өзгергіштік генетикалық материалдың (генотиптің) өзгеруінен болады, ұрпақтан –ұрпаққа беріліп отырады. Генотиптік өзгергiштiк комбинативтiк және мутациялық деп екi топқа бөледi. Комбинативтiк өзгергiштiк генотиптік өзгергіштің бір түрі, бұл жағдайда бір түрге жататын ағзалар бір бірінен генотиптегi гендердiң басқаша топтасуының нәтижесінде ажыратылады. Бұл ата анасына тән емес, жаңа белгілердің пайда болуына алып келеді. Мысалы, ата-аналарының қан топтары 2 және 3 болса, онда баласының қан тобы 4 болуы мүмкін. Комбинативтік өзгергіштіктің механизмдері: 1. Гаметогенез кезінде, мейоздың анафазасында гомологты аталық және аналық хромосомалардың тәуелсіз ажырауы; 2. Кроссинговердің нәтижесiнде генетикалық материалдың рекомбинациясы; 3. Ұрықтану кезiнде гаметалардың кездейсоқ қосылуы. Комбинативтік өзгергіштіктің биологиялық маңызы өте зор, себебі ол бір түрге жататын даралардың генетикалық әртүрлілігін, оның өкілдерінің тірі қалуын қамтамасыз етеді. Мутациялық өзгергіштік – жасушаның генетикалық аппаратына сыртқы және ішкі факторлар әсер етуінен пайда болатын өзгергіштік. Бұл жағдайда генотип те, фенотип те өзгереді. Мутация нәтижесінде пайда болған белгілер ұрпақтан ұрпаққа беріледі. Мутациялардың пайда болуы мен қалыптасу үрдісі мутагенез деп аталады. Жаңа мутациялардың пайда болуына алып келетін факторлар мутагенді факторлар деп аталады. Мутагендік факторлар физикалық (электро – магнитік сәулелер, қысым, температура, иондаушы сәулелер, радиоактивті өнімдер, УК-сәулелер және т.б.), химиялық (алкалоидтар, нитраттар, нитриттер, пестицидтер, еріткіштер, тотықтырғыштар, ауыр металл тұздары, цитостатиктер, спирттер, фенолдар және т.б.) және биологиялық (вирустар) деп жіктеледі. Адамдардаң жас мөлшері спонтанды мутацияның жиілігін арттыратын факторлардың бірі болып есептеледі. Мутациялардың көптеген жіктелулері бар:                1.Летальды 2. Зиянды 3. Нейтральды 4. Пайдалы 1.Генеративті 2. Сомалық 1.Биохимиялық 2. Физиологиялық 3. Морфологиялық |