11-органы чувств. Ш. система органов чувств общая характеристика органов чувств
 Скачать 0.92 Mb. Скачать 0.92 Mb.
|

|
Ш. СИСТЕМА ОРГАНОВ ЧУВСТВ ОБЩАЯ ХАРАКТЕРИСТИКА ОРГАНОВ ЧУВСТВ В центральную нервную систему непрерывным потоком устремляются бесчисленные нервные импульсы, обусловленные разнообразными воздействиями на организм внешней среды и постоянными изменениями, происходящими во всех его органах и тканях. Эти импульсы зарождаются в специальных приборах, называемых органами чувств, или рецепторами, служащими, по И. П. Павлову, анализаторами как внешней, так и внутренней среды организма, поэтому их и разделяют на две основные группы: экстероре-цепторы и интерорецепторы. Экстерорецепторы получают раздражения из внешней среды—химические (через органы вкуса и обоняния) и физические (через органы зрения, слуха, равновесия, осязания, терморецепторы и т. п.). Отличительная черта экстерорецепторов заключается в том, что все вызываемые ими ощущения осознаются (у человека). Интерорецепторы воспринимают раздражения с внутренних органов, сосудов, тканей. Через их посредство осуществляются: местная регуляция кровоснабжения тканей и обмена веществ; координация функций отдельных частей какой-либо системы органов; согласование деятельности различных систем организма; сигнализация в центральную нервную систему о состоянии и деятельности тех органов, в которых они расположены, и обо всех происходящих в них изменениях, как присущих норме, так и патологических. Хотя все эти импульсы в норме не доходят до сознания, тем не менее они создают общий фон для нервной деятельности в целом, как это впервые в 1886 г. подметил И М. Сеченов и назвал этот фон валовым чувством, вызывающим у человека или чувство общего благосостояния, или, напротив, чувство общего недомогания, наряду с такими общими чувствами, как голод, жажда, половое чувство, усталость или, напротив, позыв к деятельности. Особую категорию интерорецепторов составляют проприорецепторы, передающие импульсы с мускулов, сухожилий, фасций, суставов и связок и обусловливающие своеобразное суставно-мышечное чувство. При участии проприорецепторов осуществляется согласованная работа мускулов. Все указанные импульсы возникают или в свободных, или в несвободных чувствительных нервных окончаниях. Свободные нервные окончани я—это аппараты, в которых осевые цилиндры и их ветвления лежат свободно или среди клеток эпителия, не входя с ними в контакт, или в промежуточном веществе соединительной ткани (рис. 228—2,9). Они встречаются в кожном покрове, серозных оболочках, половых органах и т. п. ОБЩАЯ ХАРАКТЕРИСТИКА ОРГАНОВ ЧУВСТВ 871 Несвободные нервные окончания представляют аппараты, в которых осевые цилиндры своими ветвлениями связаны со специальными чувствительными клетками, непосредственно воспринимающими те или иные раздражения (3) (Б. И. Лаврентьев). В результате каких-то ещё неизученных процессов, происходящих в этих клетках, и рождаются импульсы в нервных волокнах.  Рис. 228. Схема строения органов чувств (по Б, И. Лаврентьеву). I—обонятельные чувствительные клетки (а); 2—свободные нервные окончания в эпителии и под эпителием; 3—вторичная чувствительная клетка с оканчивающимся на ней нервным волокном (с); 4—вкусовая луковица; 5-—диск Меркеля; 6—тельпе типа Мей-снера; 7—тельце типа Фатер—Пачини; В—хеморецептор (glomus caroticum); 9, 10, 11, 12—окончания одного и того же чувствительного нервного волокна на кровеносном сосуде и других тканях; 13*—Окончания чувствительного волокна на сосуде, эпителиальных клетках и на ганглиозных клетках; а—чувствительные клетки первичные; Ь—опорные клетки; с—нервное волокно; d—вторичная чувствительная клетка; е—капсула; /—кровеносный сосуд; 0—•мышечная ткань; ft—нервные клетки. Количество чувствительных клеток в различных рецепторах колеблется в широких пределах: то их бывает одна, как в дисках Меркеля (5), то две, как в осязательных тельцах Догеляительцах Гран-д р и, то значительное число. При этом они либо лежат среди эпителиальных клеток, будучи изолированы от них опорными клетками, как во вкусовых луковицах на языке (4), либо располагаются в соедини- 872 СИСТЕМА ОРГАНОВ ЧУВСТВ тельной ткани, будучи одеты специальными соединительнотканными капсулами в тельцах Ф а т е р —П а ч и н и (7), Гербста, Г о л ь д ж и, Маццони, Краузе. Чувствительные клетки образуют симпласт внутри капсулы в виде колбы, а в последней уже центрально располагается осевой цилиндр. В других инкапсулированных рецепторах, например в тельцах Мейснера (б), чувствительные клетки лежат пластами внутри капсулы, а между ними проходит осевой цилиндр со своими разветвлениями. Эти рецепторы отличаются друг от друга некоторыми деталями строения, различными функцией и местоположением (подробности см. в курсе гистологии), К инкапсулированным рецепторам с несвободными нервными окончаниями относятся в высшей степени сложно устроенные органы зрения истатоакустический орган у наземных животных. Несколько особняком стоит орган обоняния из чувствительных клеток, залегающих среди эпителиальных клеток обонятельной части слизистой оболочки (1). Воспринимаемые раздражения они посылают непосредственно в мозг своими отростками, формирующими в целом обонятельный нерв. У примитивных животных и органы чувств устроены примитивно и не обладают избирательностью. Они одинаково реагируют на самые различные раздражители, как физические, так и химические. Лишь в связи с усложнением в процессе эволюции взаимоотношений организма с внешней средой, а следовательно, усложнением строения и функций самого организма, возникают органы чувств своеобразного строения и функции, что обусловливает их избирательность в отношении раздражителей. Так, одни органы чувств воспринимают раздражения световой энергии, другие— от звуковых волн, третьи—химической энергии, четвёртые—различные механические раздражения. Одновременно появляются и интерорецепторы, воспринимающие раздражения, идущие из внутренних органов. Так как в примитиве раздражители действуют из внешней среды, то вполне естественно, что органы чувств впервые появляются в наружном покрове в виде первичных чувствительных клеток (рис. 152—2). Они лежат среди эпителиальных клеток, а нейриты их идут или непосредственно к исполняющему органу—мускульной клетке, или же к дендриту обособившейся нервной клетки. Первичные чувствительные клетки широко распространены у беспозвоночных и у ланцетника (рис. 230—/) у позвоночных они встречаются, повидимому, только в органах обоняния. С превращением первичных чувствительных клеток в нервные чувствительная функция их сохраняется за дендритами нервных клеток, которые как концевые, или свободные, нервные окончания разветвляются среди эпителиальных клеток кожного покрова, или под ними, или выходят на поверхность эпителия. Такие свободные нервные окончания встречаются в большом количестве у беспозвоночных. Свободные нервные окончания имеются также у позвоночных и не только в кожном покрове, но и во всех внутренних органах и тканях (рис. 228—2, 9, 11, 12, 13); они происходят из общего зачатка нервной системы и своими рецепторными отростками в процессе онтогенеза достигают периферии. С развитием из эпителиальных клеток вторичных чувствительных клеток концевые чувствительные нервные окончания вступают с ними в тесный контакт, т. е. возникают несвободные нервные окончания (3, 4, 5, 6). Вторичные чувствительные клетки имеются у некоторых беспозвоночных (у червей) и членистоногих, но закономерно они присущи только позвоночным животным. У позвоночных специальные чувствительные клетки возникают во всех органах чувств из общего зачатка нервной системы, в частности из ОБЩАЯ ХАРАКТЕРИСТИКА РАЗВИТИЯ И СТРОЕНИЯ ОРГАНОВ ЗРЕНИЯ §73 элементов глии, и, судя по исследованиям Б. И. Лаврентьева и его учеников, являются производными шванновских клеток. Такого же происхождения палочки и колбочки сетчатки глаза, а также специальные клетки статоаку-стического органа. В группе интерорецепторов различают механорецепторы, мышечные рецепторы и хеморецепторы. Проводники от этих рецепторов устремляются в центральную нервную систему через дорзальные корешки и спинномозговые ганглии. Механорецепторы сигнализируют о степени растяжения какой-либо ткани. Они характеризуются своеобразными концевыми ветвлениями нервных нитей в виде расширений или пластинок, охватывающих соединительнотканные волокна. Механорецепторы имеются всюду, но особенно много их в стенках сосудов (9, 10, 11, 12). Через мышечные рецепторы определяется степень сокращения мускулатуры, как гладкой, так и поперечнополосатой и сердечной (10). Их концевые ветвления принимают вид миниатюрных расширений или петелек. Хеморецепторы воспринимают различные изменения в крови или тканевой жидкости. Они построены по типу несвободных нервных окончаний, т. е. снабжены специальными чувствительными клетками, и образуют на сосудах особые клубочки—«гломусы» (8). К хеморецепторам относятся также параганглии и мозговое вещество надпочечников. Рецепторы внутренних органов имеют специфические особенности. Они являются «поливалентными»: одно и то же чувствительное волокно может давать одну ветвь к сосуду, а другую ветвь в гладкую мускулатуру (12), или эпителий (11), или сердечную мускулатуру (10); иногда даже третья ветвь отходит к нервной клетке межмускульного сплетения (в мышечной оболочке кишечника) (13). Этим обеспечивается передача импульса с эпителия или мышечной ткани по одному и тому же волокну (аксон-рефлекс), а одновременная связь с нервной клеткой позволяет объяснить механизм передачи раздражений с чувствительного вегетативного нейрона, не прибегая к доказательствам существования третьего парасим-патикуса (Б. И. Лаврентьев). Громадное большинство органов чувств характеризуется микроскопическим строением, поэтому в дальнейшем рассматриваются лишь органы зрения, равновесия и слуха. орган: атвнжя ОБЩАЯ ХАРАКТЕРИСТИКА РАЗВИТИЯ И СТРОЕНИЯ ОРГАНОВ ЗРЕНИЯ Чувствительность к световым раздражениям является основным свойством протоплазмы, поэтому восприятие их возможно и без помощи специальных органов, что наблюдается у простейших организмов. Обычно же у животных, реагирующих на свет, имеются специальные светочувствительные органы. Таковые встречаются даже у некоторых простейших (у евглен) в виде красного пятна, или стигмы, которое состоит из жировых капелек и красного пигмента. В примитивном виде светочувствительные органы представлены (у дождевых червей) специальными первичными чувствительными клетками с нервными отростками. Эти клетки рассеяны в кожном покрове, но попадаются также под эпидермисом или между ганглиоз-ными клетками. Более густо они расположены на головном конце, что свидетельствует о большом значении светочувствительных органов для ориентировки при движении. Простейшее усовершенствование светочувствительных органов имеет место у подвижных медуз, у которых они развиваются в виде пигментных (зрительных) пятен или ямок. Те и другие содержат скопление большого 874 СИСТЕМА ОРГАНОВ ЧУВСТВ количества светочувствительных клеток, отделённых друг от друга пигментными клетками. Последние изолируют чувствительные клетки от всестороннего действия света, а сами чувствительные клетки обращены своими рецепторными частями к свету. Ещё большее усложнение представляют так называемые обращенные, или инвертированные, глаза. Они характерны тем, что на рецепторных концах зрительных клеток формируются палочковидные отростки. Рецепторные отделы клеток обращены не к свету, как в «прямых» глазах, а в обратном направлении, и при этом погружаются  Рис. 229. Бокальчатые глазки ресничных червей. Ail—эпидермис; 2—зрительная клетка; 3—её волоски; 4—пигментная клетка; 5—её ядро. Вх 1—ядра зрительных клеток; 2—волоски клеток; 3—пигментные клетки. Стрелки 4,6,6показывают направление лучей света. в углубление (в виде чаши), образованное или одной большой пигментной клеткой, или целой группой их. Благодаря этому получается более совершенная изоляция клеток от всестороннего действия света. Органы такого строения под названием бокальчатых глазков встречаются у некоторых медуз и у ресничных червей и располагаются под эктодермой (рис. 229). Количество зрительных ямок (глазков) сильно варьирует: чем больше в них чувствительных клеток, тем меньше самих глазков (их может быть только одна пара на головном конце). Парные органы зрения становятся уже аппаратами направления, ориентирующими и контролирующими движения самого животного (в зависимости от направления света). При сращении краёв зрительных ямок возникают пузырчатые органы зрения. Уже у ряда беспозвоночных животных в пузырчатых глазах формируется светопреломляющий орган—х русталик. Он представляет собой плотное прозрачное тело то в виде двояковыпуклой чечевицы, то более шарообразной формы. Развитие хрусталика происходит или за счёт уплотнения стекловидного тела, т. е. массы, выполняющей полость глазного пузыря, или в результате утолщения эктобласта перед пузырём. Хрусталик, собирая в фокус лучи, даёт на слое зрительных клеток, или ретине (сетчатке), изображение предмета. Таким образом, с появлением хрусталика глаза из светочувствительных органов превращаются в зрительные органы, воспринимающие не только свет, тени и цвета, но также форму предметов и их расстояние. У ланцетника нет парных органов зрения; в вентральной стенке мозговой трубки, близ центрального мозгового канала, у него сосредоточено большое количество своеобразных зрительных органов—г л- а а к о в Гессе ОБЩАЯ ХАРАКТЕРИСТИКА РАЗВИТИЯ И СТРОЕНИЯ ОРГАНОВ ЗРЕНИЯ g75 (рис.230—2). Каждый глазок состоит из зрительной клетки, которая одним своим копцом погружена в чашеобразную пигментную клетку; от противоположного конца зрительной клетки отходит нейрит. Пигментные клетки обращены дорзально или вентрально. Таким образом, устроенные очень примитивно светочувствительные органы ланцетника напоминают отношения, присущие беспозвоночным животным с инвертированными глазами. Однако у ланцетника имеется и характерная особенность—тесная связь зрительных органов с нервной системой, что одновременно подчёркивает и происхождение их из эктобласта (как у беспозвоночных). У позвоночных животных парные органы зрения развиваются из стенок среднего мозгового пузыря (рис. 230—11). Эмбриональная мозговая 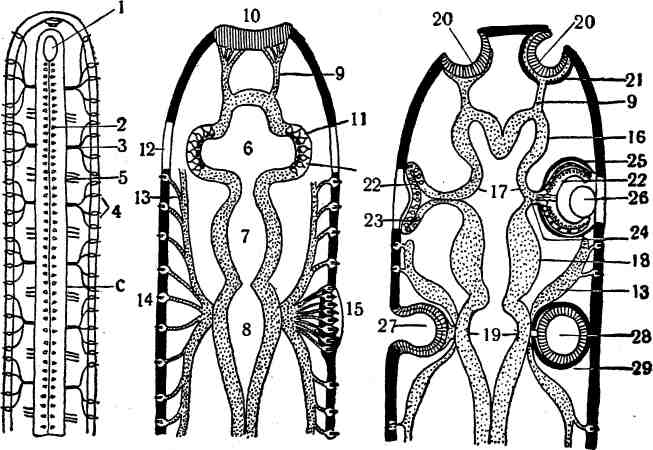 Рис. 230. Схема эволюции головного мозга и органов чувств (по А. Н. Северцову). /, II, III—последовательные стадии развития; I—головной мозг едва намечен на переднем конце спинного мозга (с); 2—глазки Гессе; 3—чувствуюпщй нерв; 4—чувствующие клетки; £—двигательный нерв; 6—передний, 7—средний и 8—задний мозг; 9—обонятельный нерв; 10—непарная обонятельная п л а кода; 11—глазной пузырь, в его стенке глазки Гессе; 12—прозрачная часть кожи против глазного пузыря; 23—нерв органа боковой линии (Л—слева), который образует слуховую плакоду {15—справа); 16—конечный, 17—промежуточный и 18—средний мозг; 19—мозжечок и продолговатый мозг; 20—парные обонятельные ямки; 21—обонятельная капсула; 22—глазной бокал с чувствительными клетками; 23—наружный пигментный слой бокала; 24—глазной нерв; 25—склера; 26—хрусталик; 27—слуховая ямка; 28—слуховой пузырек (статоцист), 29—скелетная капсула. стенка растёт к поверхности тела в виде двух глазных пузырей, которые достигают кожного покрова, а с мозгом соединяются короткими полыми ножками. В процессе дальнейшей эволюции наружная стенка пузыря вдавливается, вследствие чего последний превращается в глазной бокал (рис. 231—IV) с двойными стенками, а ножка удлиняется. Одновременно с этим с вентральной поверхности появляется впячивание и на ножке бокала для помещения артерии стекловидного тела, направляющейся к закладке хрусталика. Это вдавление образует сосудистую щель; когда края её срастаются, артерия оказывается внутри ножки и полости глазного бокала. Двойная стенка бокала диференцируется на два отдела—зрительную и слепую части. 876 СИСТЕМА ОРГАНОВ ЧУВСТВ 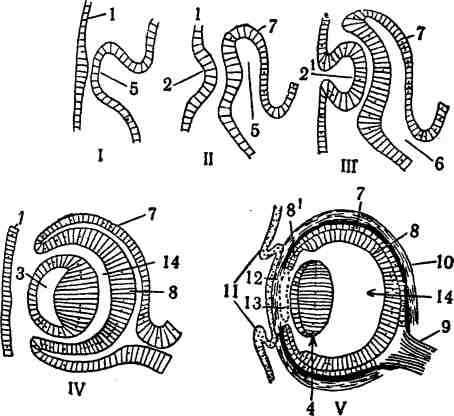 В зрительной части наружная стенка глазного бокала образует пигментный слой сетчатки (рис. 232—10; 231—7), а внутренняя стенка, разрастаясь, формирует собственно сетчатку, или ретину (рис. 231—S), состоящую из нейроэпителия (рис. 232—7, 9), прилежащего к пигментному слою, и мозгового слоя (II и III), включающего ганглиозные клетки. Мозговой слой обращен в полость бокала. Нейриты ганглиозных клеток направляются в ножку глазного бокала, которая, таким образом, превращается в зрительный нерв (рис. 231—9), вступающий в промежуточный мозг. Слепая часть глазного бокала остаётся хотя и двуслойной, но тонкостенной. Рис. 231. Схема онтогенеза глазного яблока. |