Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

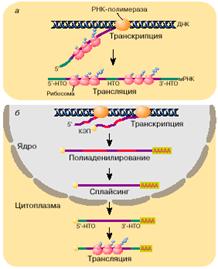
Основания и правила…Теперь посмотрим, что послужило основанием для формирования постулатов считывания ДНК, принятых в генетике: «В 1961 году Ф. Крик и С. Бреннер экспериментально показали, что делеция (вырезание) одного нуклеотида, дающая мутантный фенотип, может быть скомпенсирована вставкой нуклеотида вблизи делеции. Этот результат можно было объяснить предположив, что при делеции нуклеотида происходит сдвиг рамки считывания за местом делеции, и это приводит к полному изменению смысла всей последующей информации; при вставке одного нуклеотида вблизи места делеции происходит восстановление первоначальной рамки считывания и первоначального смысла закодированной информации. Таким образом, описанные эксперименты доказали, что генетический код не содержит запятых. В опытах с делециями и вставками Крик, Барнет, Бреннер и Уотс-Тобин (1961) также подтвердили, что код является триплетным или кратен трем, поскольку три делеции или три вставки нуклеотидов давали нормальный фенотип. Опыты Г. Виттманна по замене единичных оснований в РНК вируса табачной мозаики показали, что такие замены могут приводить к замене только одной аминокислоты в белке. Это однозначно свидетельствовало в пользу того, что генетический код не перекрывается. Другими словами, каждое основание РНК входит в состав лишь одного триплета (кодона)». [1]. То есть, если вырезать один нуклеотид … и тут же рядом вставить, то … ничего не изменится. Это подтверждает отсутствие запятых…, а так же и триплетность кода. А вот опыты Г. Виттманна - это серьезнее. Если учесть то, что сказано далее о нечувствительности трансляции к третьему элементу кода триплета, то… этот постулат не совсем верен. Неперекрываемость генетического кода не подтверждается. Коды могут перекрываться, но мы это далеко не всегда видим. Таким образом, подтвержденными можно считать только триплетность и непрерывность кода ДНК и мРНК. Вот теперь можно считывать информацию с ДНК и мРНК. Здесь правила примерно одинаковы. При этом: «…первый же встреченный на иРНК кодон AUG (Met) задает фазу последующего считывания троек, то есть служит той самой фиксированной точкой, с которой начинается считывание. Любой последующий AUG просто кодирует Met. В конце гена обязательно стоит UAA, или UAG, или UGA, а то и два нонсенса подряд». [2] Видимо, это обычный порядок считывания. Начало и конец считывания установлен. Оказывается, мы давно знаем это. Стандартная мРНК. Как мы уже говорили, процесс получения стандартной мРНК достаточно длинный. Вот как это происходит: «Эукариотические мРНК … Их транскрипция и трансляция пространственно разобщены. Транскрипция протекает в ядре, а трансляция - в цитоплазме (рис. 2, б ). Эукариотические мРНК синтезируются в виде предшественников и проходят в своем биогенезе стадию довольно сложного созревания, или процессинга. Процессинг включает в себя: 1) кэпирование 5'-конца, заключающееся в присоединении к этому концу мРНК так называемой шапочки (кэп-структуры), 2) полиаденилирование 3'-конца и, наконец, 3) сплайсинг - вырезание протяженных внутренних участков мРНК, так называемых интронов, и ковалентное воссоединение оставшихся фрагментов (экзонов) через обычную фосфодиэфирную связь…». [1].
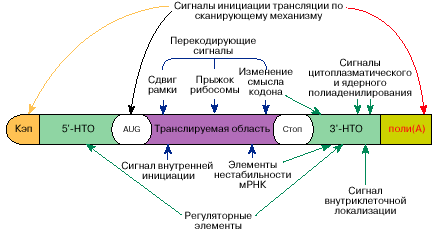
Процессинг имеет целью сформировать различные области мРНК в нужном для трансляции порядке. «Те части молекулы мРНК, в которых закодированы белки, называются транслируемыми областями. Однако помимо транслируемых областей в мРНК имеются достаточно протяженные последовательности, не кодирующие белок. Общая длина этих нетранслируемых областей порой может достигать или даже превышать длину транслируемых областей. [1] Перестановки и удаления фрагментов мРНК выполняются на этапе сплайсинга: «Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании информационной РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК)[1] и других некодирующих РНК». [8] Нетранслируемые области внутри мРНК, как мы поняли, называются интронами. Попробуем выяснить, что из себя представляют интроны, которые удалятся при сплайсинге стандартной мРНК: «Существуют две альтернативные теории, объясняющие происхождение и эволюцию сплайсосомных интронов: так называемые теории ранних интронов (РИ) и поздних интронов (ПИ). Теория РИ утверждает, что многочисленные интроны присутствовали в общих предках эу- и прокариот и, соответственно, интроны являются очень старыми структурами. Согласно этой модели, интроны были потеряны из генома прокариот. Также она предполагает, что ранние интроны способствовали рекомбинации экзонов, представляющих домены белков. ПИ утверждает, что интроны появились в генах относительно недавно и были инсертированы (вставлены) в геном после разделения организмов на про- и эукариоты. Эта модель основывается на наблюдении, что сплайсосомные интроны есть только у эукариот». [7] «Почти все эукариотические ядерные интроны начинаются с GU и оканчиваются AG (правило AG-GU)». [7]  Рис.3.Схема нуклеотидной последовательности пре-мРНК гена CDK4 человека. Большую часть последовательности занимают интроны (показаны серым цветом) Для прокариотов значимость интронов установлена более точно: «В прокариотических полицистронных мРНК имеются также внутренние межцистронные нетранслируемые области, располагающиеся между транслируемыми областями. Наряду с информацией о последовательности аминокислот в белке молекулы мРНК содержат информацию, определяющую их поведение в клетке (активность и время жизни, внутриклеточное распределение). Эта информация находится в основном в нетранслируемых областях мРНК». [1]. Вот так. Что такое интроны – узнали мало, но выяснилось много нового… Если рассматривать мРНК-предшественник, как общую информацию о том, или ином белке, то экзон надо рассматривать, как исполнительную программу производства белка на рибосоме, а интрон – дополнительная информация об этом. Например, что с чем сшивать и в каком порядке, как настроить рибосому на этот процесс, перечень дополнительных команд, и т.д. Интроны, как раз и есть кодированная запись команд и синхронизации всего производства белка - главное в этом процессе. Очень возможно, что интроны обрабатываются и частично вставляются в области 5' и 3', как дополнительная информация для считывающего устройства рибосомы. В сформированной для трансляции мРНК: Нетранслируемые области находятся на обоих концах молекул мРНК и соответственно называются 5'- и 3'-НТО». [1] Полиаденилирование 3'-конца закодировано в самой мРНК. Потому, что: «Около половины мРНК эукариот полиаденилируются на 3'-конце во время процессинга в клеточном ядре. Сигналом ядерного полиаденилирования 3'-конца служит последовательность AAUAAA, расположенная за 10-20 нуклеотидов от этого конца». [1]. «Такой последовательностью у амфибий является (U)6AUAAAG. Поли(А)-хвост на мРНК узнается особым поли(А)-связывающим белком, который участвует в инициации трансляции мРНК по кэп-зависимому механизму». [1]. И получилась мРНК, пригодная для сборки белка. Наша - такая же, как на рис 2.(б). Теперь посмотрим на рис.4. Это и есть стандартная форма мРНК. Ничего не напоминает? А ведь весьма похоже на строку из памяти компьютера. Составляющие почти те же… Только чуть сложнее. Разрешающий код, потом код начала строки и программа действий, «старт-код», собственно информационная часть, «стоп-код», проверочные и контрольные суммы, дополнительная информация о порядке выполнения действий, и, наконец, код окончания строки.
«Зрелые эукариотические мРНК, как правило, моноцистронны и кодируют только одну полипептидную цепь». [1] Таким образом, можно сказать, что стандартизация мРНК развивалась вместе с развитием клетки. |