Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

Актиновые филаменты (микрофиламенты) Порядка 7 нм в диаметре, микрофиламенты представляют собой две цепочки из мономеров актина, закрученные спиралью. В основном они сконцентрированы у внешней мембраны клетки, так как отвечают за форму клетки и способны образовывать выступы на поверхности клетки (псевдоподии и микроворсинки). Также они участвуют в межклеточном взаимодействии (образовании адгезивных контактов), передаче сигналов и, вместе с миозином — в мышечном сокращении. С помощью цитоплазматических миозинов по микрофиламентам может осуществляться везикулярный транспорт.Промежуточные филаментыДиаметр промежуточных филаментов составляет от 8 до 11 нанометров. Они состоят из разного рода субъединиц и являются наименее динамичной частью цитоскелета. Микротрубочки представляют собой полые цилиндры порядка 25 нм диаметром, стенки которых составлены из 13 протофиламентов, каждый из которых представляет линейный полимер из димера белка тубулина. Димер состоит из двух субъединиц — альфа- и бета- формы тубулина. Микротрубочки — крайне динамичные структуры, потребляющие ГТФ в процессе полимеризации. Они играют ключевую роль во внутриклеточном транспорте (служат «рельсами», по которым перемещаются молекулярные моторы — кинезин и динеин), образуют основу аксонемы ундилиподий и веретено деления при митозе и мейозе. Цитоскелет прокариот Долгое время считалось, что цитоскелетом обладают только эукариоты. с в 2001 году описывающей роль бактериальных гомологов актина в клетках Bacillus subtilis, начался период активного изучения элементов бактериального цитоскелета. К настоящему времени найдены бактериальные гомологи всех трех типов элементов цитоскелета эукариот — тубулина, актина и промежуточных филаментов[1]. Также было установлено, что как минимум одна группа белков бактериального цитоскелета. Вопрос №44.сократительные структуры в клетке. Цитоскеле́т — это клеточный каркас или скелет, находящийся в цитоплазме живой клетки. Он присутствует во всех клетках как у эукариот, так и у прокариот. Это динамичная, изменяющаяся структура, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный транспорт и клеточное деление. Кератиновые промежуточные филаменты в клетке. Цитоскелет образован белками. В цитоскелете выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических исследованиях (микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиновая система, кератины, тубулин-динеиновая система). Микротрубочки – тончайшие трубочки диаметром 24 нм, стенки которых обра-зованы белком тубулином. Глобулярные субъединицы этого белка располагаются по спирали. Микротрубочки определяют направление перемещения внутриклеточных компонентов, в том числе расхождение хромосом к полюсам клетки при делении ядра. Они участвуют в образовании «цитоскелета». Микрофиламенты – тонкие белковые нити диаметром 6 нм, состоят из белка актина, близкого тому, который содержится в мышцах. Эти нити, как и микротрубочки, являются элементами «цитоскелета». Они образуют кортикальный слой под плазматической мембраной. Кроме микротрубочек, присутствующих в цитоплазме, в клетке имеются микротрубочки, формирующие центриоли клеточного центра, базальные тельца, реснички и жгутики. Вопрос №45. Микротрубочки и их производное. Промежуточные филаменты. Микротрубочки присутствуют во всех животных клетках за исключением эритроцитов. Они образованы полимеризованными молекулами белка тубулина, который представляет собой гетеродимер, состоящий из двух субъединиц — альфа- и бета-тубулина. При полимеризации альфа-субъединица одного белка соединяется с бета-субъединицей следующего. Так формируются отдельные протофиламенты, которые, объединяясь по 13, формируют полую микротрубочку, внешний диаметр которой составляет около 25 нм, а внутренний — 15 нм. Каждая микротрубочка имеет растущий плюс-конец и медленно-растущий минус-конец. Микротрубочки — один из наиболее динамичных элементов цитоскелета. Во время наращивания длины микротрубочки присоединение тубулинов происходит на растущем плюс-конце. Разборка микротрубочек наиболее часто происходит с обоих концов. Белок тубулин, формирующий микротрубочки, не является сократительным белком, и микротрубочки не наделены способностью к сокращению и передвижению. Однако микротрубочки цитоскелета принимают активное участие в транспорте клеточных органелл, секреторных пузырьков и вакуолей. Из препаратов микротрубочек отростков нейронов (аксонов) были выделены два белка — кинезин и динеин. Одним концом молекулы этих белков ассоциированы с микротрубочкой, другим — способны связываться с мембранами органелл и внутриклеточных везикул. С помощью кинезина осуществляется внутриклеточный транспорт к плюс-концу микротрубочки, а с помощью динеина — в обратном направлении. Промежуточные филаменты (ПФ) — нитевидные структуры из особых белков, один из трех основных компонентов цитоскелета клеток эукариот. Содержатся как в цитоплазме, так и в ядре большинства эукариотических клеток. Средний диаметр ПФ — около 10 нм (9-11 нм), меньше, чем у микротрубочек (около 25 нм) и больше, чем у актиновых микрофиламентов (5-9 нм). Название получили из-за того, что толщина цитоскелетных структур, состоящих из ПФ, занимала промежуточное положение между толщиной миозиновых филаментов и микротрубочек[1]. В ядре известен только один тип ПФ — ламиновых, остальные типы — цитоплазматические. Доменная структура белковых молекул ПФ довольно консервативна. Полипептид обычно имеет два глобулярных домена на N- и C-концах, которые соединены протяженным суперскрученным палочковидным доменом, состоящим из альфа-спиралей. Основной строительный блок филамента — димер, а не мономер. Он образован двумя полипептидными цепями, обычно двух разных белков, которые взаимодействуют между собой своими палочковидными доменами, образующими двойную суперскрученную спираль. Цитоплазматические ПФ образованы из таких димеров, образующих неполярные нити, толщиной в один блок. Отсутствие полярности у ПФ обусловлено антипараллельной ориентацией димеров в тетрамере. Из них далее образуются более сложные структуры, в которых ПФ могут уплотняться, вследствие чего имеют непостоянный диаметр. В отличие от актина и тубулина белки ПФ не имеют сайта сязывания нуклеозидтрифосфатов. Цитоплазматические ПФ есть не у всех эукариот, они обнаружены только у некоторых групп животных. Так, ПФ есть у нематод. моллюсков и позвоночных. но не найдены у членистоногих и иглокожих. У позвочноных ПФ отсутствуют в некоторых клетках (например, олигодендроцитах). В растительных клетках ПФ не обнаружены.[2][3] В большинстве животных клеток ПФ образуют «корзинку» вокруг ядра, откуда направлены к периферии клеток. ПФ особенно много в клетках, подверженных механическим нагрузкам: в эпителиях, где ПФ участвуют в соединении клеток друг с другом через десмосомы, в нервных волокнах, в клетках гладкой и поперечно-полосатой мышечной ткани. Вопрос №46. Специализированные структуры плазмотической мембраны (микроворсинки, реснички, жгутики) Микроворсинка — вырост эукариотической (обычно животной) клетки, имеющий пальцевидную форму и содержащий внутри цитоскелет из актиновых микрофиламентов. Из микроворсинок состоит воротничок у клеток хоанофлагеллят и у воротничково-жгутиковых клеток губок и других многоклеточных животных. В организме человека микроворсинки имеют клетки эпителия тонкого кишечника, на которых микроворсинки формируют щеточную кайму, а также механорецепторы внутреннего уха — волосковые клетки.Микроворсинки нередко путают с ресничками, однако они резко отличаются по строению и функциям. Реснички имеют базальное тело и цитоскелет из микротрубочек, способны к быстрым движениям (кроме видоизмененных неподвижных ресничек) и служат у крупных многоклеточных обычно для создания токов жидкости или восприятия раздражителей, а у одноклеточных и мелких многоклеточных животных также для передвижения. Микроворсинки не содержат микротрубочек и способны лишь к медленным изгибаниям (в кишечнике) либо неподвижны. За упорядочение актинового цитоскелета микроворсинок отвечают вспомогательные белки, взаимодействующие с актином — фимбрин, спектрин, виллин и др. Микроворсинки также содержат цитоплазматический миозин нескольких разновидностей. Микроворсинки кишечника (не путать с многоклеточными ворсинками) во много раз увеличивают площадь поверхности всасывания. Кроме того. у позвоночных на их плазмалемме закреплены пищеварительные ферменты, обеспечивающие пристеночное пищеварение. Микроворсинки внутреннего уха (стереоцилии) интересны тем, что образуют ряды с различной, но строго определенной в каждом ряду длиной. Вершины микроворсинок более короткого ряда соединены с более длинными микроворсинками соседнего ряда с помощью белков - протокадгеринов. Их отсутствие или разрушение может приводить к глухоте, так как они необходимы для открывания натриевых каналов на мембране волосковых клеток и, следовательно, для преобразования механической энергии звука в нервный импульс [1] Хотя микроворсинки сохраняются на волосковых клетках в течение всей жизни, каждая из них постоянно обновляется за счет тредмиллинга актиновых филаментов, Реснички — органеллы, представляющие собой тонкие (диаметром 0,1—0,6 мкм) волосковидные структуры на поверхности эукариотических клеток. Длина их может составлять от 3—15 мкм до 2 мм (реснички гребных пластинок гребневиков). Могут быть подвижны или нет: неподвижные реснички играют роль рецепторов. Характерны для инфузорий. У многих беспозвоночных животных ими покрыта вся поверхность тела (ресничные черви, личинки кишечнополостных и губок) или отдельные его участки (например, жабры у полихет и двустворчатых моллюсков, подошва ноги у брюхоногих моллюсков[источник не указан 362 дня]). У коловраток из специализированных ресничек состоит коловращательный аппарат. У многих беспозвоночных (кишечнополостные, гребневики, турбеллярии и др.) реснички также имеются на клетках кишечного эпителия. У позвоночных (в том числе человека) клетки с подвижными ресничками также есть во многих органах. У человека ресничным эпителием выстланы дыхательные пути, евстахиевы трубы, семявыносящие канальцы, желудочки мозга и спинномозговой (центральный) канал[источник не указан 362 дня]. Видоизмененные реснички служат световоспринимающим аппаратом фоторецепторов сетчатки глаза и воспринимающим запахи аппаратом хеморецепторов обонятельного эпителия. Снаружи покрыты мембраной, являющейся продолжением плазмолеммы — цитоплазматической мембраны. В центре проходит две полные (состоящие из 13 протофиламентов) микротрубочки, на периферии — девять пар микротрубочек, из которых в каждой паре одна полная, а вторая неполная (состоит из 11 протофиламентов). У основания находится базальное тело (кинетосома), имеющее в поперечном разрезе ту же структуру, что и половинка центриоли, то есть состоящее из девяти троек микротрубочек. К каждой полной микротрубочке периферических пар (дублетов) вдоль всей её длины присоединены «ручки» из двигательного белка динеина (см. статью аксонема). При гидролизе АТФ головки динеина «шагают» по микротрубочке соседнего дублета. Если бы микротрубочки не были закреплены на кинетосоме, это вызвало бы скольжение дублетов друг относительно друга. Такое скольжение наблюдается в эксперименте на ресничках, обработанных трипсином (длина аксонемы при добавлении АТФ увеличивается в результате в 9 раз). В интактной ресничке происходит изгибание дублетов и, в результате, всей реснички. Как правило, реснички совершают удары в одной плоскости. У инфузорий прямой удар (продвигающий клетку вперед) ресничка совершает в выпрямленном состоянии, а возвратный — в изогнутом. Как регулируется согласованное изгибание разных дублетов, видимо, неизвестно. При деполяризации мембраны и поступлении внутрь клетки ионов кальция у инфузорий направление прямого удара может меняться на противоположное Жгутик — поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. Жгутики прокариот и эукариот резко различаются: бактериальный жгутик имеет толщину 10—20 нм и длину 3—15 мкм, он пассивно вращается расположенным в мембране мотором; жгутики же эукариот толщиной до 200 нм и длиной до 200 мкм, они могут самостоятельно изгибаться по всей длине. У эукариот часто также присутствуют реснички, идентичные по своему строению жгутику, но более короткие (до 10 мкм). Вопрос №47. Мышечные волокна как надклеточные структуры. Как известно, все ткани организма имеют клеточную структуру, не представляют исключение и мышцы. Поэтому мне придется провести краткий экскурс в цитологию – науку о клетке, и напомнить читателям о роли и свойствах основных структур клетки. В грубом приближении клетка состоит из двух важнейших, взаимосвязанных между собой частей – цитоплазмы и ядра. Ядро – содержит в себе молекулы ДНК , в которых заключена вся наследственная информация. ДНК – полимер, закрученный в виде двойной спирали, каждая спираль которого составлена из огромного количества четырех видов мономеров, называемых нуклеотидами. Последовательность нуклеотидов в цепочке кодирует все белки организма. Ядро ответственно за размножение клетки – деление . Деление клетки начинается с разделения молекулы ДНК на две спирали, каждая из которых способна достроить парную из набора свободных нуклеотидов и вновь превратится в молекулу ДНК. Таким образом, количество ДНК в ядре удваивается, далее ядро делится на две части, а за ним и вся клетка. Цитоплазма – это все, что в клетке окружает ядро. Она состоит из цитозоли (клеточной жидкости), в которую включены различные органеллы, такие как митохондрии, лизосомы, рибосомы и прочие. Митохондрии – это энергетические станции клетки, в них с помощью различных ферментов происходит окисление углеводов и жирных кислот. Энергия, выделяющаяся при окислении веществ, идет на присоединение третьей фосфатной группы к молекуле Аденезиндифосфата (АДФ) с образованием Аденезинтрифосфата (АТФ) – универсального источника энергии для всех процессов, протекающих в клетке. Отсоединяя третью фосфатную группу и вновь превращаясь в АДФ, АТФ выделяет запасенную ранее энергию. Ферменты или Энзимы – вещества белковой природы в сотни и тысячи раз увеличивающие скорость протекания химических реакций. Практически все жизненно важные химические процессы в организме происходят только в присутствии специфических ферментов. Лизосомы – округлые пузырьки, содержащие около 50 ферментов. Лизосомные ферменты расщепляют поглощенный клеткой материал и собственные внутренние структуры клетки (автолизис). Лизосомы, сливаясь в фагосомы, способны переваривать целые органеллы, подлежащие дезинтеграции. Рибосомы – органеллы, на которых происходит сборка белковой молекулы. Клеточная мембрана – оболочка клетки, она обладает избирательной проницаемостью, то есть способностью пропускать одни вещества и задерживать другие. Задача мембраны сохранять постоянство внутренней среды клетки. СТРОЕНИЕ МЫШЦЫ.Структурно функциональной единицей скелетной мышцы является симпласт или мышечное волокно – огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (в дальнейшем под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект). Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. Мышечное волокно, как и любая клетка, окружено оболочкой – сарколемой . Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна. Группы мышечных волокон образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани, переходящей на концах мышцы в сухожилия, крепящиеся к кости. 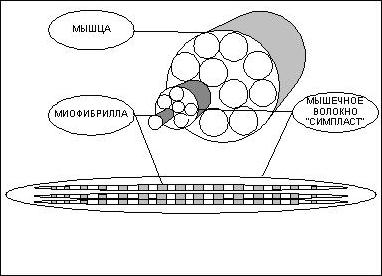 Вопрос №48. Роль рибосом в клетке. Рибосома — важнейший немембранный органоид живой клетки сферической или слегка эллипсоидной формы, диаметром 100—200 ангстрем, состоящий из большой и малой субъединиц. Рибосомы служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией. В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Нередко с одной молекулой мРНК ассоциировано несколько рибосом, такая структура называется полирибосомой. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре — ядрышке. Рибосомы представляют собой нуклеопротеид, в составе которого отношение РНК/белок составляет 1:1 у высших животных и 60-65:35-40 у бактерий. Рибосомная РНК составляет около 70 % всей РНК клетки. Рибосомы эукариот включают четыре молекулы рРНК, из них 18S, 5.8S и 28S рРНК синтезируются в ядрышке РНК полимеразой I в виде единого предшественника (45S), который затем подвергается модификациям и нарезанию. 5S рРНК синтезируется РНК полимеразой III в другой части генома и не нуждаются в дополнительных модификациях. Почти вся рРНК находится в виде магниевой соли, что необходимо для поддержания структуры; при удалении ионов магния рибосома подвергается диссоциации на субъединицы. Константа седиментации (скорость оседания в ультрацентрифуге) рибосом эукариотических клеток равняется 80S (большая и малая субъединицы 60S и 40S, соответственно), бактериальных клеток (а также митохондрий и пластид) — 70S (большая и малая субъединицы 50S и 30S, соответственно). Трансляция — синтез белка рибосомой на основе информации, записанной в матричной РНК (мРНК). мРНК связывается с малой субъединицей рибосомы, когда происходит узнавание 3'-концом 16S рибосомной РНК комплементарной последовательности Шайн-Далгарно, расположенной на 5'-конце мРНК (у прокариот), а также позиционирование стартового кодона (как правило, AUG) мРНК на малой субъединице. У эукариот малая субчастица рибосомы связывается также с помощью кэпа, на конце мРНК. Ассоциация малой и большой субъединиц происходит при связывании формилметионил-тРНК (fMET-тРНК) и участии факторов инициации (IF1, IF2 и IF3 у прокариот; их аналоги и дополнительные факторы участвуют в инициации трансляции у эукариотических рибосом). Таким образом, распознавание антикодона (в тРНК) происходит на малой субъединице. После ассоциации, fMET-тРНК находится в P- (peptidyl-) сайте каталитического(пептидил-трансферазного) центра рибосомы. Следующая тРНК, несущая на 3'-конце аминокислоту и комплементарная второму кодону на мРНК, помещается с помощью фактора EF-Tu в А- (aminoacyl-) сайт каталитического центра рибосомы. Затем, образуется пептидная связь между формилметионином (связанным с тРНК, находящейся в Р-сайте) и аминокислотой, принесенной тРНК, находящейся в А-сайте. Механизм катализа образования пептидной связи в пептидил-трансферазном центре до сих пор полностью не ясен. На данный момент существует несколько гипотез, объясняющих детали этого процесса: 1. Оптимальное позиционирование субстратов (induced fit)[5], 2. Исключение из активного центра воды, способной прервать образование пептидной цепи посредством гидролиза [6], 3. Участие нуклеотидов рРНК (таких как А2450 и А2451) в переносе протона[7][8], 4. Участие 2'-гидроксильной группы 3'-концевого нуклеотида тРНК (А76) в переносе протона [9];. Высокая эффективность катализа достигается взаимодействием этих факторов. Вопрос №49. Образоваине и роль рибосом в клетке. Рибосомы — цитоплазматические органеллы, на которых происходит синтез белка. Рибосомы могут функционировать только в комплексе с двумя другими типами РНК — транспортной РНК, доставляющей аминокислоты к строящейся молекуле белка, и матричной РНК, служащей источником информации, необходимой для сборки заданной последовательности аминокислот. Таким образом, рибосому можно сравнить с мастерской по производству белковых молекул. Образование рибосом в ядрышках. Гены, отвечающие за синтез рибосомной РНК, располагаются в пяти парах хромосом и представлены в виде множества копий, что позволяет одновременно синтезировать большое количество рибосомной РНК, необходимой для реализации клеточных функций. Сформировавшиеся рибосомы накапливаются в ядрышках— специализированных структурах ядра, связанных с хромосомами. Если клетка синтезирует много белка, в ней образуется большое количество рибосомной РНК, поэтому ядрышки в этой клетке крупные. Напротив, в клетках, синтезирующих мало белка, ядрышки бывают даже не видны. Рибосомная РНК в ядрышках связывается с рибосомными белками с образованием глобулярных частиц, представляющих собой отдельные субъединицы рибосомы. Эти субъединицы отделяются от ядрышка, выходят из ядра через поры ядерной мембраны и распределяются почти по всей цитоплазме. Попав в цитоплазму, субъединицы собираются в зрелую функционирующую рибосому. Зрелых рибосом в ядре нет, поэтому синтез белка осуществляется только в цитоплазме клетки. Роль рибосом: служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией Вопрос 50:Морфология ядерных структур.
Вопрос 51. Роль ядерных структур в жизнедеятельности клетки Приведенный в главе 2 краткий обзор основных процессов, связанных с синтезом белка, в принципе одинаковых у всех форм живого, указывает на особое значение клеточного ядра. Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую - с ее реализацией, с обеспечением синтеза белка. В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее, в ядре происходит воспроизведение или редупликация и разъединение (сегрегация) молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном и количественном смысле объемы генетической информации. В ядре эукариот происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток. Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственного аппарата белкового синтеза. Это не только синтез, транскрипция, на молекулах ДНК разных информационных РНК, но также транскрипция всех видов трансферных РНК и рибосомных РНК. В ядрах эукариотических клеток происходит «созревание» (процессинг, сплайсинг) первичных транскриптов. В ядре эукариот происходит также образование субъединиц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро. Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал воспроизводится и функционирует. Поэтому выпадение или нарушение любой из перечисленных выше функций гибельно для клетки в целом. Так, нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что тоже гибельно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке ли к грубым его нарушениям. Все это указывает на ведущее значение ядерных структур в процессах, связанных с синтезом нуклеиновых кислот и белков - основных функционеров в жизнедеятельности клетки. Однако, что необходимо еще раз подчеркнуть, что функционирование ядра как системы хранения и реализации генетической информации сопряжено, неразрывно связано, с другими функциональными системами клетки, которые обеспечивают работу ядра специальными белками, потоком предшественников, энергией и пр. Вопсро 52. Структура ядрышка. Ядрышко - источник рибосом. Строение рибосом. Амплификация ядрышек. Внутри интерфазных ядер как при витальных наблюдениях, так и на фиксированных и окрашенных препаратах видны мелкие, обычно шаровидные тельца - ядрышки. В живых клетках они выделяются на фоне диффузной организации хроматина. Ядрышки являются наиболее плотными структурами в клетке. Ядрышки обнаруживаются практически во всех ядрах эукариотических клеток. Это говорит об обязательном присутствии этого компонента в клеточном ядре. В клеточном цикле ядрышко присутствует в течение всей интерфазы: в профазе по мере компактизации хромосом во время митоза оно постепенно исчезает, и отсутствует в мета- и анафазе, и вновь появляется в середине телофазы, чтобы сохраняться вплоть до следующего митоза, или до гибели клетки. Ядрышки представлялись как структурное выражение хромосомной активности. Ядрышки содержат РНК, стала понятна их «базофилия», сродство к основным красителям, из-за кислой природы РНК. По данным цитохимических и биохимических исследований основным компонентом ядрышка является белок: на его долю приходится до 70-80% от сухого веса. Такое большое содержание белка и определяет высокую плотность ядрышек. Кроме белка в составе ядрышка обнаружены были нуклеиновые кислоты: РНК (5-14%) и ДНК (2-12%). Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Все они построены в клетке одинаково, имеют одинаковую молекулярную композицию, выполняют одинаковую функцию - синтез белка - поэтому их можно так же считать клеточными органоидами. В отличие от других органоидов цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.) они представлены в клетке огромным числом: за клеточный цикл их образуется 1 х 107 штук. Поэтому основная масса клеточной РНК представляет собой именно рибосомную РНК. РНК рибосом относительно стабильна, рибосомы могут существовать в клетках культуры ткани в течение нескольких клеточных циклов. В печеночных клетках время полужизни рибосом составляет 50-120 часов. Рибосомы - это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных (неповторенных) белков и несколько молекул РНК, Рибосомы прокариот и эукариот по своим размерам и молекулярным характеристикам отличаются, хотя и обладают общими принципами организации и функционирования. К настоящему времени методом рентгеноструктурного анализа высокого разрешения полностью расшифрована структура рибосом. Амплифицированные ядрышки - гены рРНК мб избыточно реплицированы. При этом дополнительная репликация генов рРНК происходит в целях обеспечения продукции большого количества рибосом. В результате такого сверхсинтеза генов рРНК их копии могут становиться свободными, экстрахромосомными. Эти внехромосомные копии генов рРНК могут функционировать независимо, в результате чего возникает масса свободных дополнительных ядрышек, но уже не связанных структурно с ядрышкообразующими хромосомами. Это явление получило название амплификации генов рРНК. подробно изучено на растущих ооцитах амфибий. У X. laevis амплификация рДНК, происходит в профазеI. В этом случае количество амплифицированной рДНК (или генов рРНК) становится в 3000 раз больше того, что приходится на гаплоидное количество рДНК, и соответствует 1,5х106 генов рРНК. Эти сверхчисленные внехромосомные копии и образуют сотни дополнительных ядрышек в растущих ооцитах. В среднем же на одно дополнительное ядрышко приходится несколько сот или тысяч генов рРНК. Амплифицированные ядрышки встречаются также в ооцитах насекомых. У окаймленного плавунца в ооцитах обнаружено 3х106 экстрахромосомных копий генов рРНК. После периода созревания ооцита при его двух последовательных делениях дополнит ядрышки в состав митотических хромосом не входят, они отделяются от новых ядер и деградируют. У Tetrachymena pyriformis в гаплоидном геноме микронуклеуса единственный ген рРНК. В макронуклеусе 200 копий. У дрожжей экстрахромосомные копии генов рРНК - циклические ДНК l3 мкм, сод один ген рРНК. Вопрос 53. Ядро-система хранения, воспроизведение и реализации генетического материала. Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм. Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой. Наружная ядерная (1) мембрана со стороны, обращенной в цитоплазму, покрыта рибосомами, придающими ей шероховатость, внутренняя (2) мембрана гладкая. Ядерные мембраны являются частью мембранной системы клетки: выросты наружной ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов. Кариоплазма (ядерный сок, нуклеоплазма) — внутреннее содержимое ядра, в котором располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды. Ядрышко (4) представляет собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и варьирует от 1 до 7 и более. Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают. Ядрышко образуется на определенных участках хромосом, несущих информацию о структуре рРНК. Такие участки называются ядрышковым организатором и содержат многочисленные копии генов, кодирующих рРНК. Из рРНК и белков, поступающих из цитоплазмы, формируются субъединицы рибосом. Таким образом, ядрышко представляет собой скопление рРНК и рибосомальных субъединиц на разных этапах их формирования Вопрос 54. Организация и свойства клеточного ядра. Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК). В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на молекуле ДНК. В ядре же синтезированные молекулы РНК подвергаются ряду модификаций, после чего выходят в цитоплазму. Образование субъединиц рибосом также происходит в ядре в специальных образованиях - ядрышках. Клеточное ядро имеет важнейшее значение в жизнедеятельности клетки, поскольку служит хранилищем наследственной информации, содержащейся в хромосомах. Ядро ограничено ядерной оболочкой, отделяющей его содержимое (кариоплазму) от цитоплазмы. Оболочка состоит из двух мембран, разделенных промежутком. Обе они пронизаны многочисленными порами, благодаря которым возможен обмен веществами между ядром и цитоплазмой. В ядре клетки у большинства эукариот находится от 1 до 7 ядрышек. С ними связаны процессы синтеза РНК и тРНК. Основные компоненты ядра – хромосомы, образованные из молекулы ДНК и различных белков. В световом микроскопе они хорошо различимы лишь в период клеточного деления (митоза, мейоза). В неделящейся клетке хромосомы имеют вид длинных тонких нитей, распределенных по всему объему ядра. Во время деления клеток хромосомные нити образуют плотные спирали, вследствие чего становятся видимыми (с помощью обычного микроскопа) в форме палочек, «шпилек». Весь объем генетической информации распределен между хромосомами ядра. В процессе их изучения были выявлены следующие закономерности: ♦ в ядрах соматических клеток (т. е. клеток тела, неполовых) у всех особей одного вида содержится одинаковое количество хромосом, составляющих набор хромосом (рис. 19); ♦ для каждого вида характерен свой хромосомный набор по их количеству (например, у человека 46 хромосом, у мушки дрозофилы – 8, у аскариды – 4, у речного рака – 196, у лошади – 66, у кукурузы – 104); ♦ хромосомы в ядрах соматических клеток могут быть сгруппированы парами, получившими название гомологичных хромосом на основании их сходства (по строению и функциям); ♦ в ядрах половых клеток (гамет) из каждой пары гомологичных хромосом содержится только одна, т. е. общий набор хромосом вдвое меньше, чем в соматических клетках; ♦ одинарный набор хромосом в половых клетках называется гаплоидным и обозначается буквой n, а в соматических – диплоидным (2n). Из изложенного ясно, что каждая пара гомологичных хромосом образована объединением отцовских и материнских хромосом при оплодотворении, т. е. слиянии половых клеток (гамет). И наоборот, при образовании половых клеток из каждой пары гомологичных хромосом в гамету попадает только одна. В теле хромосом выделяют первичную перетяжку (называемую центромерой), к которой прикрепляются нити веретена деления. Она делит хромосому на два плеча. Хромосомы могут быть равноплечими, разноплечими и одноплечими. Вопрос №55.Опишите роль ядрав реализации генетической информации. Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК). В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на молекуле ДНК. В ядре же синтезированные молекулы РНК подвергаются ряду модификаций, после чего выходят в цитоплазму. Образование субъединиц рибосом также происходит в ядре в специальных образованиях - ядрышках. Клеточное ядро имеет важнейшее значение в жизнедеятельности клетки, поскольку служит хранилищем наследственной информации, содержащейся в хромосомах. Ядро ограничено ядерной оболочкой, отделяющей его содержимое (кариоплазму) от цитоплазмы. Оболочка состоит из двух мембран, разделенных промежутком. Обе они пронизаны многочисленными порами, благодаря которым возможен обмен веществами между ядром и цитоплазмой. В ядре клетки у большинства эукариот находится от 1 до 7 ядрышек. С ними связаны процессы синтеза РНК и тРНК. Основные компоненты ядра – хромосомы, образованные из молекулы ДНК и различных белков. В световом микроскопе они хорошо различимы лишь в период клеточного деления (митоза, мейоза). В неделящейся клетке хромосомы имеют вид длинных тонких нитей, распределенных по всему объему ядра. Во время деления клеток хромосомные нити образуют плотные спирали, вследствие чего становятся видимыми (с помощью обычного микроскопа) в форме палочек, «шпилек». Весь объем генетической информации распределен между хромосомами ядра. В процессе их изучения были выявлены следующие закономерности: ♦ в ядрах соматических клеток (т. е. клеток тела, неполовых) у всех особей одного вида содержится одинаковое количество хромосом, составляющих набор хромосом (рис. 19); ♦ для каждого вида характерен свой хромосомный набор по их количеству (например, у человека 46 хромосом, у мушки дрозофилы – 8, у аскариды – 4, у речного рака – 196, у лошади – 66, у кукурузы – 104); ♦ хромосомы в ядрах соматических клеток могут быть сгруппированы парами, получившими название гомологичных хромосом на основании их сходства (по строению и функциям); ♦ в ядрах половых клеток (гамет) из каждой пары гомологичных хромосом содержится только одна, т. е. общий набор хромосом вдвое меньше, чем в соматических клетках; ♦ одинарный набор хромосом в половых клетках называется гаплоидным и обозначается буквой n, а в соматических – диплоидным (2n). Из изложенного ясно, что каждая пара гомологичных хромосом образована объединением отцовских и материнских хромосом при оплодотворении, т. е. слиянии половых клеток (гамет). И наоборот, при образовании половых клеток из каждой пары гомологичных хромосом в гамету попадает только одна. В теле хромосом выделяют первичную перетяжку (называемую центромерой), к которой прикрепляются нити веретена деления. Она делит хромосому на два плеча. Хромосомы могут быть равноплечими, разноплечими и одноплечими. Вопрос №56. Поток информации в клетке. |