Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

|
Митохондрия (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) — двумембранная гранулярная или нитевидная органелла толщиной около 0,5 мкм. Характерна для большинства эукариотических клеток как аутотрофов (фотосинтезирующие растения), так и гетеротрофов (грибы, животные). Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии в синтезе молекул АТФ, который происходит за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300000 и 500000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. Происхождение митохондрий: в соответствии с теорией симбиогенеза, митохондрии появились в результате того, что примитивные клетки (прокариоты), которые не могли сами использовать кислород для генерирования энергии и что накладывало серьёзные ограничения на возможности развития, захватывали бактерии (прогеноты), которые могли это делать. В процессе развития таких отношений прогеноты передали множество своих генов сформировавшемуся, благодаря повысившейся энергоэффективности, ядру теперь уже эукариот.[1] Вот почему современные митохондрии больше не являются самостоятельными организмами. Хотя их геном кодирует компоненты собственной системы синтеза белка, многие ферменты и белки, необходимые для их функции, кодируются хромосомами, синтезируются в клетке и только потом транспортируются в органеллы. Число митохондрий в клетке непостоянно. Их особенно много в клетках, в которых потребность в кислороде велика. По своему строению они представляют собой цилиндрические органеллы, встречающиеся в эукариотической клетке в количестве от нескольких сот до 1-2 тысяч и занимающие 10-20 % её внутреннего объёма. Размеры от 1 до 70 мкм и форма митохондрий. При этом ширина этих органелл относительно постоянна 0,5-1 мкм. Способны изменять форму. В зависимости от того, в каких участках клетки в каждый конкретный момент происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры цитоскелета. Альтернативой множеству разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих АТФ небольшие участки цитоплазмы, является существование длинных и разветвлённых митохондрий, каждая из которых может энергетически обеспечивать отдалённые друг от друга участки клетки. Вариантом такой протяжённой системы может также являться упорядоченное пространственное объединение множества митохондрий, обеспечивающее их кооперативную работу и встречающееся как у одноклеточных, так и у многоклеточных организмов. Особенно сложно этот тип хондриома устроен в скелетных мышцах млекопитающих, где группы гигантских разветвлённых митохондрий связаны друг с другом с помощью межмитохондриальных контактов (ММК). Последние образованы плотно прилегающими друг к другу наружными митохондриальными мембранами, в результате чего межмембранное пространство в этой зоне имеет повышенную электронную плотность. Особенно обильно ММК представлены в клетках сердечных мышц, где они связывают множественные отдельные митохондрии в согласованную работающую кооперативную систему. Альтернативой множеству разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих АТФ небольшие участки цитоплазмы, является существование длинных и разветвлённых митохондрий, каждая из которых может энергетически обеспечивать отдалённые друг от друга участки клетки. Вопрос №31. Образование АТФ в митохондриях. Ионизация водорода, цепь транспорта электронов, образование воды. Первым этапом окислительного фосфорилирования в митохондриях является ионизация атомов водорода, извлекаемых из питательных веществ. Как было изложено ранее, атомы водорода извлекаются парами; при этом один атом незамедлительно становится ионом водорода (Н+), а другой объединяется с НАД+, образуя НАД-Н. В верхней части рисунка показана последовательность событий, происходящих с НАД-Н и Н+. Начальный этап состоит в отщеплении атома водорода от НАД-Н и образовании при этом еще одного иона водорода. В итоге воссоздается НАД+, который может быть вновь использован. Электроны отделяются от атомов водорода, в результате водород ионизируется. Электроны сразу поступают в цепь переноса электронов, являющуюся неотъемлемой частью внутренней мембраны (мембраны, снабженной криптами) митохондрий, к акцепторам электронов. Акцепторы электронов могут обратимо восстанавливаться и окисляться, присоединяя и отдавая электроны. Важным компонентом цепи переноса электронов являются флавопротеины, некоторые протеинаты сульфида железа, убихинон цитохромы В,С1, С, А и А3. Каждый электрон передается от одного акцептора к другому до тех пор, пока не достигнет цитохрома А3, названного цитохромоксидазой за его способность передавать два электрона кислороду и таким образом превращать его в ионизированную форму, которая может образовывать воду путем взаимодействия с ионами водорода. На рисунке показан перенос электронов по дыхательной цепи с окончательным использованием цитохромоксидазой кислорода с образованием молекул воды. Перенос электронов по дыхательной цепи сопровождается выделением энергии, используемой для синтеза АТФ следующим образом. Закачивание ионов водорода в наружную камеру митохондрий, обеспечиваемое цепью переноса электронов. В процессе прохождении электронов по цепи переноса электронов выделяется большое количество энергии. Эта энергия используется для перекачивания ионов водорода от внутреннего матрикса митохондрий в наружную камеру, расположенную между внутренней и наружной мембранами (левая часть рисунка). Это создает высокую концентрацию положительно заряженных ионов водорода в пространстве между внутренней и наружной мембранами. В то же время формируется высокий отрицательный потенциал матрикса митохондрии. Образование АТФ. Следующий этап окислительного фосфорилирования состоит в превращении АДФ в АТФ. Эта возможность опосредована существованием крупных белковых молекул, расположенных на поверхности внутренней мембраны митохондрий. Прободая ее насквозь, головки молекул выдаются в матрикс. Эти молекулы представляют собой АТФ-азу. Ее называют АТФ-синтетазой. Высокая концентрация положительно заряженных ионов водорода в наружной камере и высокая разность потенциалов по обе стороны внутренней мембраны вызывают движение ионов водорода во внутренний матрикс митохондрии через молекулу АТФ-азы. Выделяемая при таком продвижении ионов водорода энергия используется АТФ-азой для превращения АДФ в АТФ путем объединения АДФ со свободным фосфатным радикалом (P1). Так формируется вторая макроэргическая фосфатная связь в молекуле. Окончательным этапом этого процесса является перевод АТФ из матрикса митохондрии наружу в цитоплазму клетки. Этот процесс осуществляется путем облегченной диффузии через внутреннюю мембрану и путем простой диффузии через проницаемую наружную мембрану митохондрии. В свою очередь, АДФ, постоянно движущийся в противоположном направлении, превращается в АТФ. На каждые два электрона, проходящих до конца по цепи переноса электронов (полученных путем ионизации 2 атомов водорода), приходится 3 молекулы синтезируемой АТФ. Вопрос №32. Организзация генома митохондрий. ДНК в митохондриях представлена циклическими молекулами, не образующими связь с гистонами, в этом отношении они напоминают бактериальные хромосомы. У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представлены множественными копиями, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре. Так же как и у бактерий митохондральная ДНК собрана в отдельную зону – нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток. При слиянии митохондрий может происходить обмен их внутренними компонентами. рРНК и рибосомы митохондрий резко отличны от таковых в цитоплазме. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s). В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий. На митохондриальном геноме синтезируются и транспортные РНК, всего синтезируется 22 тРНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х105. В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х106. Вопрос №33. Гладкий ретикулум. Строение и футнкции. Эндоплазматическая сеть (эндоплазматичеческий ретикулум) — органоид клетки эукариот. В виде сети каналов и цистерн, ограниченных одинарной мембраной, она разветвлена по всему объему цитоплазмы. Эндоплазматическая сеть участвует в обмене веществ: синтезирует липиды для наружной двойной мембраны клетки и для собственной, одинарной мембраны, обеспечивает транспорт веществ между органоидами клетки, служит копилкой веществ и местом их изоляции. Различают два типа эндоплазматической сети — шероховатую и гладкую. Шероховатая сеть несет на наружной поверхности многочисленные рибосомы. Функции: синтезированные на них белки здесь изолируются от других белков клетки путем переноса их через мембрану канала эндоплазматической сети. Она «узнает» пропускаемые белки по их особым «сигнальным» концам. Отщепление этих концов после прохождения белка через мембрану называют созреванием белка. Одни белки — секреторные — выделяются из клетки. Другие включаются во все мембраны клетки. Гладкая сеть состоит из трубочек, каналов и пузырьков меньшего сечения, чем в шероховатой сети. Ее функции так же разнообразны: здесь синтезируются липиды мембран, но, кроме них, и немембранные липиды (например, особые гормоны животных), специальными ферментными комплексами обезвреживаются ядовитые вещества, накапливаются ионы. Так, в поперечнополосатых мышцах гладкая сеть служит резервуаром ионов кальция. Мембраны этой сети содержат мощные кальциевые «насосы», которые в сотые доли секунды переносят в любую сторону большое количество ионов кальция. В специализированных клетках вид гладкой сети различен, что связано с ее конкретными функциями во внутриклеточном обмене. Эндоплазматическая сеть очень ранима при воздействиях: она быстро теряет рибосомы и разрушается. Однако благодаря способности к быстрым перестройкам может восстанавливаться. Функции гладкой эндоплазматической сети:
Пластинчатый комплекс Гольджи (сетчатый аппарат) представлен скоплением уплощенных цистерн и небольших везикул, ограниченных билипидной мембраной. Пластинчатый комплекс подразделяется на субъединицы - диктиосомы. Каждая диктиосома представляет собой стопку уплощенных цистерн, по периферии которых локализуются мелкие пузырьки. При этом, в каждой уплощенной цистерне периферическая часть несколько расширена, а центральная сужена. Вопрос №34. Строение и функции гранулярного ретикула Эндоплазматическая сеть (эндоплазматичеческий ретикулум) — органоид клетки эукариот. В виде сети каналов и цистерн, ограниченных одинарной мембраной, она разветвлена по всему объему цитоплазмы. Эндоплазматическая сеть участвует в обмене веществ: синтезирует липиды для наружной двойной мембраны клетки и для собственной, одинарной мембраны, обеспечивает транспорт веществ между органоидами клетки, служит копилкой веществ и местом их изоляции. Различают два типа эндоплазматической сети — шероховатую и гладкую. Шероховатая сеть несет на наружной поверхности многочисленные рибосомы. Функции: синтезированные на них белки здесь изолируются от других белков клетки путем переноса их через мембрану канала эндоплазматической сети. Она «узнает» пропускаемые белки по их особым «сигнальным» концам. Отщепление этих концов после прохождения белка через мембрану называют созреванием белка. Одни белки — секреторные — выделяются из клетки. Другие включаются во все мембраны клетки. Гладкая сеть состоит из трубочек, каналов и пузырьков меньшего сечения, чем в шероховатой сети. Ее функции так же разнообразны: здесь синтезируются липиды мембран, но, кроме них, и немембранные липиды (например, особые гормоны животных), специальными ферментными комплексами обезвреживаются ядовитые вещества, накапливаются ионы. Так, в поперечнополосатых мышцах гладкая сеть служит резервуаром ионов кальция. Мембраны этой сети содержат мощные кальциевые «насосы», которые в сотые доли секунды переносят в любую сторону большое количество ионов кальция. В специализированных клетках вид гладкой сети различен, что связано с ее конкретными функциями во внутриклеточном обмене. Эндоплазматическая сеть очень ранима при воздействиях: она быстро теряет рибосомы и разрушается. Однако благодаря способности к быстрым перестройкам может восстанавливаться. Функции гладкой эндоплазматической сети:
Пластинчатый комплекс Гольджи (сетчатый аппарат) представлен скоплением уплощенных цистерн и небольших везикул, ограниченных билипидной мембраной. Пластинчатый комплекс подразделяется на субъединицы - диктиосомы. Каждая диктиосома представляет собой стопку уплощенных цистерн, по периферии которых локализуются мелкие пузырьки. При этом, в каждой уплощенной цистерне периферическая часть несколько расширена, а центральная сужена. Билет 35. Строение аппарата Гольджи. Секреторная функция аппарата Гольджи. Комплекс Гольджи представляет собой стопку мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. Практически все секретируемые клеткой вещества (как белковой, так и небелковой природы) проходят через аппарат Гольджи и там упаковываются в секреторные пузырьки. Мембранные элементы АГ участвуют в сегрегации и накоплении продуктов, синтезированных в ЭР, участвуют в их химических перестройках, созревании: это, главным образом перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран. В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов. Но главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, АГ является источником клеточных лизосом. Участие АГ в процессах выведения секреторных продуктов было очень хорошо изучено на примере экзокринных клеток поджелудочной железы. Для этих клеток характерно наличие большого числа секреторных гранул (зимогеновых гранул), которые представляют собой мембранные пузырьки, заполненные белковым содержимым. В составе белков зимогеновых гранул входят разнообразные ферменты: протеазы, липазы, карбогидразы, нуклеазы. При секреции содержимое этих зимогеновых гранул выбрасывается из клеток в просвет железы, а затем перетекает в полость кишечника. Так как основным продуктом, выводимым клетками поджелудочной железы, является белок, то исследовали последовательность включения радиоактивных аминокислот в различные участки клетки. Для этого животным вводили меченную тритием аминокислоту (3Н-лейцин) и с помощью электронно-микроскопической радиоавтографии следили во времени за локализацией метки. Оказалось, что через короткий промежуток времени (3-5 мин) метка локализовалась только в базальных участках клеток, в участка, богатых гранулярным ЭР. Так как метка включалась в белковую цепь во время синтеза белка, то было ясно, что ни в зоне АГ, ни в самих зимогеновых гранулах синтез белка не происходит, а он синтезируется исключительно в эргастоплазме на рибосомах. Несколько позднее (через 20-40 мин) метка кроме эргастоплазмы была обнаружена в зоне вакуолей АГ. Следовательно, после синтеза в эргастоплазме белок был транспортирован в зону АГ. Еще позднее (через 60 мин) метка обнаруживалась уже и в зоне зимогеновых гранул. В дальнейшем метку можно было видеть в просвете ацинусов этой железы. Таким образом, стало ясно, что АГ является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Также подробно процессы синтеза и выведения белков были изучены на других клетках (молочная железа, бокаловидные клетки кишечника, щитовидная железа и др.), и были исследованы морфологические особенности этого процесса. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ. По мере модификации белков в цистернах аппарата Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз. Такое описание событий является только общей схемой участия аппарата Гольджи в секреторных процессах. дело усложняется тем, что одна и та же клетка может участвовать в синтезе многих выделяемых белков, может их друг от друга изолировать и направлять к клеточной поверхности или же в состав лизосом. В аппарате Гольджи происходит не просто "перекачка" продуктов из одной полости в другую, но и постепенно идет их "созревание", модификация белков, которая заканчивается "сортировкой" продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям. Билет 36.модификация белков в аппарате Гольджи. Сортировка белков в АГ В цистернах аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам органеллы, в которых происходит их модификации — гликозилирование и фосфорилирование. При О-гликозилировании к белкам присоединяются сложные сахара через атом кислорода. При фосфорилировании происходит присоединение к белкам остатка ортофосфорной кислоты. Разные цистерны аппарата Гольджи содержат разные резидентные каталитические ферменты и, следовательно, с созревающими белками в них последовательно происходят разные процессы. Понятно, что такой ступенчатый процесс должен как-то контролироваться. Действительно, созревающие белки «маркируются» специальными полисахаридными остатками (преимущественно маннозными), по-видимому, играющими роль своеобразного «знака качества». Существуют две взаимонеисключающие гипотезы, объясняющие этот механизм:
Вопрос 37. Лизосомы. Образование строение функция. гетерогенность лизосом. Патологии лизосом. Лизосома — клеточный органоид размером 0,2 — 0,4 мкм, один из видов везикул. Эти одномембранные органоиды — часть вакуома (эндомембранной системы клетки). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты. Функциями лизосом являются:
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером болезней накопления может служить амавротическая идиотия при накоплении гликогена.
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи. Лизосомы — гетерогенные органеллы, имеющие разную форму, размеры, ультраструктурные и цитохимические особенности. «Типичные» лизосомы животных клеток обычно имеют размеры 0,1-1 мкм, сферическую или овальную форму. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных). Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путем фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
Вопрос 38. Опишите путь секреторного белка от места синтеза белка до выхода из клетки. В клетках, в которых секреция происходит в ответ на внеклеточный сигнал , секретируемые белки концентрируются и хранятся в секреторных пузырьках (их часто называют секреторными гранулами из-за темной сердцевины). При получении соответствующего сигнала они высвобождаются путем экзоцитоза . Секреторные пузырьки отпочковываются от транс-сети Гольджи . Полагают, что для их образования нужен клатрин и связанные с ним белки, создающие "кайму", потому что часть поверхности формирующихся пузырьков обычно покрыта клатрином. Эта кайма удаляется вскоре после того, как пузырек полностью сформируется ( рис.8-76 ). Подобно лизосомным гидролазам белки, предназначенные для секреторных пузырьков (их часто называют секреторными белками ) должны быть отобраны и упакованы в соответствующие пузырьки в транс-сети Гольджи . По-видимому, в этом случае происходит избирательная агрегация секреторных белков. Образовавшиеся агрегаты в электронном микроскопе выглядят как электроноплотный материал в транс-сети Гольджи. "Сигнал сортировки" , направляющий белок к таким агрегатам, неизвестен, но видимо, это сигнальный участок, общий для многих секреторных белков. Такой вывод подтверждается следующими данными: если ген, кодирующий секреторный белок, перенести в секреторную клетку другого типа, в норме не синтезирующую данный белок, то чужой белок будет также упаковываться в секреторные пузырьки. Неизвестно, каким образом при образовании секреторных пузырьков отбираются агрегаты, содержащие секреторные белки. Секреторные пузырьки имеют уникальные мембранные белки , часть из которых может служить рецепторами (в транс-сети Гольджи ) для связывания агрегированного материала, подлежащего упаковке. Секреторные пузырьки больше транспортных пузырьков, переносящих лизосомные гидролазы , и агрегаты, которые в них содержатся, слишком велики для того, чтобы каждая молекула секретируемого белка могла связаться с рецептором в мембране пузырька , как это происходит при транспорте лизосомных ферментов . Захват этих агрегатов секреторными гранулами скорее напоминает поглощение частиц при фагоцитозе на клеточной поверхности, которое также происходит с участием покрытых клатрином мембран . После того, как незрелые секреторные пузырьки отпочкуются от транс- сети Гольджи, они утрачивают кайму, и их содержимое сильно концентрируется. Такая конденсация происходит резко и, возможно, вызывается закислением среды в полости пузырька за счет работы ATP- зависимой протонной помпы в его мембране. Агрегация секретируемых белков (или других компонентов) и последующая их конденсация в секреторных пузырьках обусловливает увеличение концентрации этих белков в 200 раз по сравнению с аппаратом Гольджи . Благодаря этому секреторные пузырьки имеют возможность высвобождать по "команде" большие количества материала. Вопрос №39.Опишите путь гидролаз от места их синтеза до места назначения. ГИДРОЛАЗЫ, класс ферментов, катализирующих гидролиз. Могут действовать на сложноэфирные и гликозидные связи, на связи С—О в простых эфирах. С—S в сульфидах, С—N в пет идах, и др. Гидролазы, катализирующие гидролиз сложноэфирных связей (эстеразы), действуют на сложные эфиры карбоновых и тио-карбоновых кислот, моноэфиры фосфорной кислоты и др. К этому подклассу относятся, в частности, ферменты, играющие важную роль в метаболизме липидов. нуклеиновых кислот и нуклеозидов. например арилсульфатазы, ацетилхолинэстераза, дезоксирибонуклеазы. липазы, фосфатазы, фосфолипазыи эндодезоксирибонуклеазы Ферменты, катализирующие гидролиз связи С—N в пептидах и белках (пептидгидролазы),- самая многочисленная группа гидролазы К ним относятся ферменты, отщепляющие одну или две аминокислоты с N- или С-конца полипептидной цепи (напр., аминопептидазы, карбоксипептидазы), а также эндопептидазы, или протеиназы, расщепляющие цепь вдали от концевых остатков. Пептидгидролазы играют важную роль не только в катаболизме белков и пептидов, но и в биол. регуляции (гормональной регуляции, активации проферментов, регуляции кровяного давления и солевого обмена и т.д.). Вопрос 40. Опишите путь макромолекулы от момента поступления её в клетку до момента усвоения. знаю Вопрос 41. Роль АГ и ЭР в регенерации и обновлениях поверхностного аппарата клетки (ПАК) Роль АГ в обновлении ПАК: Аппарат Гольджи. Во многих клетках животных, например в нервных, он имеет форму сложной сети, расположенной вокруг ядра. В клетках рас-тений и простейших аппарат Гольджи представлен отдельными тельцами серповидной или палочковидной формы. Строение этого органоида сходно в клетках растительных и животных организмов, несмотря на разнообразие его формы. В состав аппарата Гольджи входят: полости, ограниченные мембранами и расположенные группами (по 5-10); крупные и мелкие пузырьки, расположенные на концах полостей . Все эти элементы составляют единый комплекс. Аппарат Гольджи выполняет много важных функций. По каналам эндо-плазматической сети к нему транспортируются продукты синтетической деятельности клетки - белки, углеводы и жиры. Все эти вещества сна-чала накапливаются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму и либо используются в самой клетке в процессе ее жизнедеятельности, либо выводятся из нее и используются в организме. Например, в клетках поджелудочной железы млекопитающих синтезируются пищеварительные ферменты, которые накапливаются в полостях органоида. Затем образуются пузырьки, наполненные ферментами. Они выводятся из клеток в проток поджелудочной железы, откуда перетекают в полость кишечника. Еще одна важная функция этого органоида заключается в том, что на его мембранах происходит синтез жиров и углеводов (полисахаридов), которые используются в клетке и которые входят в состав мембран. Благодаря деятельности аппарата Гольджи происходят обновление и рост плазматической мем-браны. (см. ниже и про АГ и про ЭР, 2-а других источника). Роль ЭР в обновлении ПАК: Эндоплазматический ретикулум (эндоплазматическая сеть) представляет собой систему цистерн, канальцев и вакуолей, ограниченных цитомембраной. Различают гранулярный (шероховатый) и агранулярный (гладкий) эндоплазматический ретикулум; в первом преобладают плоские мешочки — цистерны, во втором — канальцы. Мембраны шероховатого ретикулума со стороны гиалоплазмы покрыты рибосомами. Степень развития этого органоида зависит от уровня метаболической активности и дифференцировки К.: он сильнее развит в клетках, активно синтезирующих белки. (ещё один источник). ЭР – транспорт белков. Полость ЭР отделяется от цитозоля одиночной мембраной ( мембраной ЭР ), служащей связующим звеном между этими двумя компартментами. Наоборот полости ЭР и каждой цистерны аппарата Гольджи отделены друг от друга двумя мембранами и цитозолем, поэтому транспорт макромолекул между этими органеллами осуществляется при помощи транспортных пузырьков . Все вновь синтезированные белки, независимо от их места назначения (полость ЭР, аппарат Гольджи, лизосомы или внеклеточное пространство) сначала поступают в полость ЭР. Некоторые белки переходят из цитозоля в шероховатый ЭР сразу после их синтеза. Это белки двух типов: 1) трансмембранные , которые лишь частично переносятся через мембрану ЭР и остаются заключенными в нее, и 2) водорастворимые , которые полностью переносятся через мембрану ЭР и освождаются в его полость. В клетках млекопитающих импорт белков в ЭР начинается еще до того, как полипептидная цепь полностью синтезирована, т. е. он происходит одновременно с трансляцией (котрансляционно). Таким образом, в цитоплазме имеется две пространственно изолированные популяции рибосом. Одни из них ( рибосомы, связанные с мембраной ), расположены на обращенной к цитоплазме поверхности мембраны ЭР и заняты синтезом белков, которые сразу же переносятся внутрь ЭР. Другие ( рибосомы свободные ) не приклеплены ни к какой мембране и производят все остальные белки, кодируемые ядром. Связанные и свободные рибосомы идентичны по строению и функции. Они различаются только по белкам, которые синтезируются на них в каждый данный момент. Если рибосоме достается синтез белка с сигнальным пептидом для ЭР, то такой сигнал направляет рибосому к мембране ЭР. (ещё один источник). Мы уже подчеркивали, насколько обширны структуры эндоплазматического ретикулума и аппарата Гольджи в секреторных клетках. В основе этих структур лежат мембраны из липидных бислоев, сходные по строению с мембраной клетки. Стенки мембран содержат ферменты, которые катализируют синтез многих веществ, необходимых клетке. Большая часть синтетических процессов происходит в эндоплазматическом ретикулуме. Сформированные здесь вещества направляются в аппарат Гольджи, где они перед выходом в цитоплазму подвергаются дальнейшей обработке. Вначале следует остановиться на веществах, которые синтезируются в отдельных областях ретикулума и аппарата Гольджи. Синтез белков на шероховатом эндоплазматическом ретикулуме. На наружной поверхности шероховатого эндоплазматического ретикулума содержится большое количество прикрепленных к нему рибосом; на них происходит синтез белка, незначительное количество которого попадает в цитозоль, а основная часть — в просвет трубочек и пузырьков ретикулума, т.е. в эндоплазматический матрикс. Синтез липидов в гладком эндоплазматическом ретикулуме. Эндоплазматический ретикулум способен к синтезу липидов, особенно фосфолипидов и холестерола. Они быстро растворяются в мембранном бислое, что способствует дальнейшему разрастанию структур ретикулума, в основном гладкого. Небольшие пузырьки, называемые транспортными, или ЭР-вакуолямиу постоянно отделяются от мембран гладкого ретикулума, предотвращая таким образом его избыточный рост. Большая часть этих транспортных вакуолей затем быстро направляется в аппарат Гольджи. Другие функции эндоплазматического ретикулума. Эндоплазматический ретикулум, особенно гладкий, обладает и другими важными функциями. 1. Обеспечение ферментами, расщепляющими гликоген при необходимости получения из него энергии. 2. Обеспечение большим количеством ферментов, способных нейтрализовать вредные для клетки вещества, например лекарственные препараты. Способы обезвреживания включают коагуляцию, окисление, гидролиз, соединение с глюкуроновой кислотой и т.п. 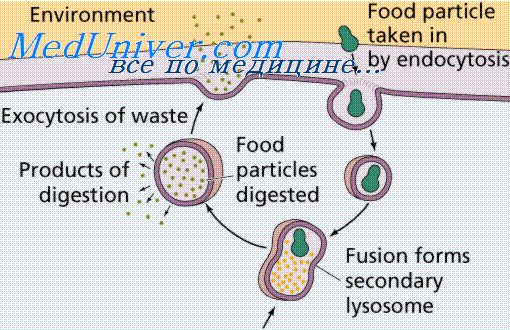 |