Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

Поток информацииЖизнедеятельность клетки как единицы биологической активности обеспечивается совокупностью взаимосвязанных, приуроченных к определенным внутриклеточным структурам, упорядоченных во времени и пространстве обменных (метаболических) процессов. Эти процессы образуют три потока: информации, энергии и веществ. Благодаря наличию потока информации клетка на основе многовекового эволюционного опыта предков приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, а также передает в ряду поколений. В потоке информации участвуют ядро (конкретно ДНК хромосом), макромолекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). На завершающем этапе этого потока полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуры и используются в качестве катализаторов или структурных белков (рис. 2.7). Кроме основного по объему заключенной информации ядерного генома в эукариотических клетках функционируют также геномы митохондрий, а в зеленых растениях — и хлоропластов. 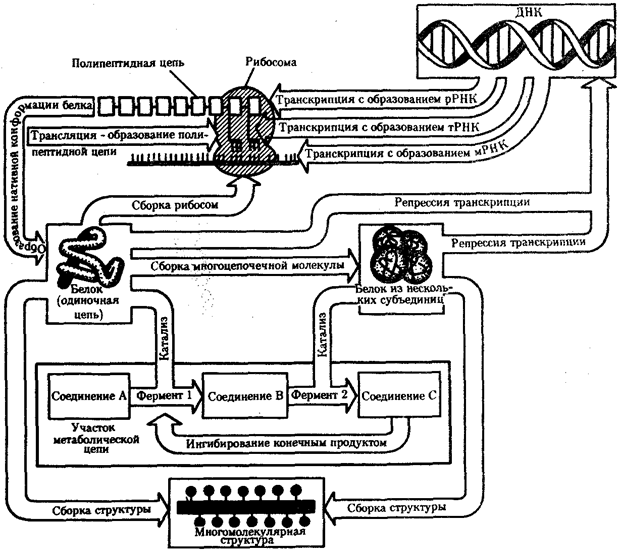 Вопрос 57. Организация эу- и гетерохроматина. Структура и химия хромасаомы. Гетерохроматин – неактивная, компактно упакованная разновидность ядерного вещества – хроматина. В хромосомах гетерохроматиновые участки расположены преимущественно в районах центромер. По сравнению с эухроматиновыми участками они содержат меньше структурных генов (кодирующих белки или полипептиды), отличаются более поздней репликацией (удвоением). Расположение гетерохроматина на хромосомах очень индивидуально. Основные отличия гетерохроматина от эухроматина. 1) Плотная упаковка ДНК. В топологически разомкнутой ДНК не может существовать торсионное напряжение. Различия в упаковке эу- и гетерохроматиновых блоков метафазных хромосом выявляются не только при окраске по Гимза (G- и С-полосы), но также при использовании электронного или фазово-контрастного микроскопа. По завершении митоза компактное состояние ДНК частично сохраняется и проявляется в виде хромоцентров интерфазного ядра. Плотная упаковка ДНК деконденсируется холодом и колцемидом. 2) Поздняя репликация ДНК. В настоящее время принято считать, что это не запаздывание репликации ДНК гетерохроматина в S-периоде клеточного цикла, а просто ДНК гетерохроматина имеет другое расписания репликации. 3) Подавлена транскрипция (инактивация Х хромосомы?). В качестве доказательства приводится пример: ядра покоящихся лимфоцитов периферической крови почти полностью гетерохроматизированы и транскрипционно неактивны. 4) Гетерохроматин вызывает эффект положения гена. В рамках обсуждаемой гипотезы автор утверждает, что ген, попавший в блок гетерохроматина, должен быть инактивирован, потому что разрывы в его ДНК будут индуцированы одновременно с индукцией разрывов в молекулах ДНК всего блока в процессе его гетерохроматинизации. 5) Склонность к агрегации (липкость). Неоднократно отмечалась упорядоченность хромосом в митозе, приписываемая «притяжению» между гетерохроматиновыми районами гомологов. Описана конъюгация гетерохроматиновых районов сестринских хроматид [2, 3]. В политенных ядрах нередко образуются эктопические контакты между блоками интеркалярного гетерохроматина разных хромосом [4]. Все эти проявления липкости могут быть следствием образования гибридных ДНК, включающих в себя однонитевые участки двух разных блоков гетерохроматина. 6) Высокая частота локализации концов перестроек хромосом. Общепринято, что хромосомная перестройка получается после разрыва пары внутри- или межхромосомных связей, инверсии концов и их последующего восстановления. Автор гипотезы предполагает, что наличие однонитевых разрывов с «липкими» концами в гетерохроматиновой ДНК заметно облегчает образование гибридных участков и тем самым увеличивает частоту перестроек, концы которых оказываются локализованными в блоках гетерохроматина 7) Формируется в онтогенезе через разрывы ДНК. Процесс формирования гетерохроматина в раннем развитии животных достаточно продолжителен и сопряжен с разрывами ДНК. У самок мышей гетерохроматинизация Х-хромосомы происходит в те же сроки, и тем же механизмом, что и формирование гетерохроматина. 8) Вариабельность количества ДНК. 9) Гетерохроматин обогащен повторами и мобильными генетическими элементами. 10) Подавлена рекомбинация в мейозе. Вопрос 58. Уровни структурной организации хроматина Сохраняя преемственность в ряду клеточных поколений, хроматин в зависимости от периода и фазы клеточного цикла меняет свою организацию. В интерфазе при световой микроскопии он выявляется в виде глыбок, рассеянных в нуклеоплазме ядра. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец — хромосом. Интерфазную и метафазную формы существования хроматина расценивают как два полярных варианта его структурной организации, связанных в митотическом цикле взаимопереходами. В пользу такой оценки свидетельствуют данные электронной микроскопии о том, что в основе как интерфазной, так и метафазной формы лежит одна и та же элементарная нитчатая структура. В процессе электронно-микроскопических и физико-химических исследований в составе интерфазного хроматина и метафазных хромосом были выявлены нити (фибриллы) диаметром 3,0—5,0, 10, 20—30 нм. Полезно вспомнить, что диаметр двойной спирали ДНК составляет примерно 2 нм, диаметр нитчатой структуры интерфазного хроматина равен 100—200, а диаметр одной из сестринских хроматид метафазной хромосомы — 500— 600 нм. Наиболее распространенной является точка зрения, согласно которой хроматин (хромосома) представляет собой спирализованную нить. При этом выделяется несколько уровней спирализации (компак-тизации) хроматина (табл. 3.2).
Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки. Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом (рис. 3.46, Б). В связи с этим геном человека, состоящий из 3 · 109 п. н., представлен двойной спиралью ДНК, упакованной в 1,5 · 107 нуклеосом. Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм.
Интерфазная хромонема. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы (рис. 3.48). Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки Хроматиновая фибрилла диаметром 20—30 нм преобразуется в структуру диаметром 100—200 нм, называемую интерфазной хромонемой. Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Вопрос 59. Первый уровень компактизации ДНК. Структурная роль нуклиосом. Нуклиосомы при репликации. Политенные хромосомы. В ранних биохимических и электронномикроскопических работах было показано, что препараты ДНП содержат нитчатые структуры с диаметром от 5 до 50 нм. Постепенно стало ясно, что диаметр фибрилл хроматина зависит от способа выделения препарата. На ультратонких срезах интерфазных ядер и митотических хромосом после фиксации глутаровым альдегидом обнаруживались хроматированные фибриллы толщиной 30 нм. Такие же размеры имели фибриллы хроматина при физической фиксации ядер - при быстром замораживании ядер, скалывании объекта и получении реплик с таких препаратов. В последнем случае исключалось воздействие на хроматин переменных химических условий. Но все эти методы и приемы не давали никакой информации о характере локализации ДНК и гистонов в хроматиновых фибриллах. Крупным событием в изучении хроматина было открытие двумя разными способами нуклеосом - дискретных частиц хроматина. Так при осаждении на подложку для электронной микроскопии препаратов хроматина в щелочных условиях при низкой ионной силе, можно было видеть, что нити хроматина представляли собой что-то, напоминающее "бусы на нитке": небольшие, около 10 нм, глобулы, связанные друг с другом отрезками ДНК длиной около 20 нм. Эти наблюдения совпадали с результатами фракционирования хроматина после частичного нуклеазного переваривания. Было найдено, что если подвергнуть действию нуклеазы микрококков выделенный хроматин, то он подвергается распаду на регулярно повторяющиеся структуры. Так ДНК, полученная из хроматина, обработанного нуклеазой, состояла из серии отрезков, кратных 200 парам оснований; встречались отрезки в 200, 400, 600, 800 и больше пар нуклеотидов (п.н.). Это говорит о том, что нуклеазной атаке в составе хроматина подвергаются участки ДНК, расположенные примерно через каждые 200 п.н. При этом в кислоторастворимую фракцию (низкополимерная) ДНК уходит всего 2% ядерной ДНК. Кроме того после такой нуклеазной обработки из хроматина путем центрифугирования удается выделить фракцию частиц со скоростью седиментации 11S (S - единица Сведберга, определяющая скорость седиментации частиц, равна 1 х 10-13 с), а также частицы кратного этой величине размера: димеры, тримеры, тетрамеры и т.д. Оказалось, что частицы 11S содержат ДНК около 200 п.н. и восемь гистонов (октамер) по две копии гистонов H2A, H2B, H3 и H4 и одну копию гистона H1. Такая сложная нуклеопротеидная частица получила название нуклеосомы. Более подробный анализ этой фракции показал, что нуклеосома устроена следующим образом: октамер гистонов образует белковую основу-сердцевину (от англ. core, часто в нашей литературе этот термин используется без перевода: кор, коровая частица), по поверхности которой располагается ДНК величиной в 146 п.н., образующая 1,75 оборота; остальные 54 п.н. ДНК образуют участок, несвязанный с белками сердцевины - линкер, который, соединяя две соседние нуклеосомы, переходит в ДНК следующей нуклеосомы. Гистон H1 связывается частично с основной, сердцевиной и с участком линкера (около 30 п.н.). Следовательно, полная нуклеосома содержит около 200 п.н. ДНК (146 п.н.- сердцевина, 30 п.н. - участок линкера в комплексе с гистоном H1, 30 п.н. - свободная ДНК), октамер сердцевинных (коровых) гистонов и одну молекулу гистона H1. Молекулярная масса полной нуклеосомы - 262000 Да. Рассчитано, что на весь гаплоидный геном человека (3 х 109 пар оснований) приходится 1,5 х 107 нуклеосом. Сердцевина или коровая частица (или минимальная нуклеосома) очень консервативны по своей структуре: они всегда содержат 146 п.н. ДНК и октамер гистонов. Линкерный участок может значительно варьировать (от 8 до 114 п.н. на нуклеосому). Используя метод рассеяния нейтронов удалось установить форму и точные размеры нуклеосом. При грубом приближении - это плоский цилиндр или шайба диаметром 11 нм и высотой 6 нм. Располагаясь на подложке для электронного микроскопирования они образуют «бусины», глобулярные образования около 10 нм, гуськом, тандемно сидящие на вытянутых молекулах ДНК. На самом же деле вытянутыми являются только линкерные участки, остальные три четверти длины ДНК спирально уложены по периферии гистонового октамера. Сам гистоновый октамер, как считают, имеет форму, напоминающую мяч для игры в рэгби, в состав которого входит тетрамер (H3 · H4)2 и два независимых димера H2A · H2B. В фибриллах хроматина линкерный участок не линеен, а продолжая спираль ДНК на поверхности нуклеосомной частицы,связывает соседние нуклеосомы так, что образуется как бы сплошная нить, толщиной около 10 нм, состоящая из тесно расположенных нуклеосом. При этом за счет дополнительной спирализации ДНК (1 отрицательный супервиток ДНК на 1 нуклеосому) происходит первичная компактизация ДНК, с плотностью упаковки равной 6-7 (200 п.н. длиной 68 нм, уложены в глобулу диаметром 10 нм). Укладка почти двух витков ДНК по периферии сердцевин нуклеосомы происходит, как считается, за счет взаимодействия положительно заряженных аминокислотных остатков на поверхности октамера гистонов с фосфатами ДНК. N- и C-концевые участки сердцевинных гистонов, обогащенные положительными зарядами, вероятно, служат для дополнительной стабилизации структуры нуклеосомы. Ведущая роль сердцевинных (коровых) белков в компактизации ДНК показана при самосборке нуклеосом. Регулируя последовательность добавления гистонов и ДНК, удалось получить полную реконструкцию нуклеосом. В этом процессе не играет никакой роли источник, откуда была взята ДНК: это может быть ДНК бактерии и даже циклическая ДНК вирусов. Оказалось, что для образования нуклеосом гистон H1 не требуется, он участвует в связывании уже готовых нуклеосом друг с другом и в образовании более высоких уровней компактизации ДНК. Ключевыми в построении нуклеосом оказались гистоны H3 и H4. При этом вначале ДНК связывается с тетрамером (H3 · H4)2 к которому позжеприсоединяются два димера H2A · H2B. Вероятно, высокая консервативность в строении гистонов H3 и H4 отражает их ведущую структурную роль на первых этапах компактизации ДНК при образовании нуклеосом. Вопрос 60. Второй и третий уровень организации хромотина. Петлевые домены ДНК - третий уровень структурной организации хроматина Расшифровка принципа строения элементарных хромосомных компонентов - нуклеосом и 30 нм фибрилл - еще мало что дает для понимания основ трехмерной организации хромосом, как в интерфазе, так и в митозе. Сорокакратное уплотнение ДНК, которое достигается при сверхспиральном характере ее компактизации, совершенно еще недостаточно для получения реального (1 х 104) уровня уплотнения ДНК. Следовательно должны существовать более высокие уровни компактизации ДНК, которые в конечном счете должны определять размеры и общие характеристики хромосом. Такие высшие уровни организации хроматина были обнаружены при искусственной его деконденсации, когда было найдено, что поддержание их связано с негистоновыми белками. В этом случае специфические белки связываются с особыми участками ДНК, которые в местах связывания образуют большие петли или домены. Как уже указывалось, сложная структура ядра или нуклеоида прокариот организована в виде иерархии петлевых доменов ДНК, связанных с небольшим количеством специальных белков.Петлевой принцип упаковки ДНК обнаруживается также и у эукариотических клеток. Так если выделенные ядра обработать 2 М NaCI, т.е. удалить все гистоны, то целостность ядра сохраняется, за исключением того, что вокруг ядра возникнет т.н. «гало», состоящее из огромного числа петель ДНК. Такая структура ядер получила название «нуклеоида». Гало состоит из огромного (до 50000) количества замкнутых на периферии петель ДНК, со средним размером петель около 60 т.п.н., основание которых закреплено где-то внутри ядра, на участках негистоновых белков. Тем самым считается, что после удаления гистонов основания петлевых доменов ДНК, связаны с т.н. «матриксом» или «скэффолдом» - негистоновым белковым остовом интерфазного ядра. Оказалось, что петлевые домены ДНК интерфазных ядер можно выделить. В выделенных ядрах в присутствии двухвалентных катионов (2 мМ Ca++ ) в хроматине ядра выявляются небольшие сгустки величиной около 100 нм, т.н. хромомеры. Если такие хромомеры препаративно выделить, а затем экстрагировать из них гистоны, то под электронным микроскопом можно видеть розетковидные петлистые структуры, где отдельные петли отходят от центрального плотного участка. Количество петель в такой розетке может составлять 15-80, а общая величина ДНК может достигать 200 т.п.н., с суммарной длиной ДНК до 50 мкм. Обработка таких розеток протеиназами приводит к исчезновению плотной центральной области розетки и к разворачиванию петель ДНК.Сходные картины можно наблюдать при разрыхлении политенных хромосом. Здесь хромомеры в виде розеток хроматина выявляются в зонах хроматиновых дисков, в то время как междисковые участки их не содержат. При деконденсации хроматина ядер некоторых растений (Allium, Haemantnus, Vicia), для которых характерна особая структура интерфазных ядер, хромомеры видны в составе хромонемных нитей.Подобные розетковидные петлистые структуры, хромомеры, можно видеть также при разрыхлении и митотических хромосом как животных, так и растений. Следовательно, хромосомные 30 нм фибриллы, состоящие из ДНК и гистонов, упаковываются в виде петлистых розетковидных структур, претерпевая еще дополнительную компактизацию. Это третий уровень структурной организации хроматина, как считается, может приводить уже к 600-кратной компактизации ДНК.Важно отметить, что размер отдельных петлевых доменов совпадает с размером среднихрепликонов и может соответствовать одному или нескольким генам. В своих основаниях петли ДНК связаны негистоновыми белками ядерного матрикса, в состав которых могут входить как ферменты репликации ДНК, так и транскрипции. Такая петельно-доменная структура хроматина обеспечивает не только структурную компактизацию хроматина, но и организует функциональные единицы хромосом - репликоны и транскрибируемые гены. Комплекс белков, участвующих в такой структурно-функциональной организации хроматина, относится к белкам ядерного матрикса. Второй уровень компактизациии - 30 нм фибрилла: Т.о первый, нуклеосомный, уровень компактизации хроматина играет как регуляторную, так и структурную роль, обеспечивая плотность упаковки ДНК приблизительно в 6-7 раз.Однако во многих электронномикроскопических исследованиях было показано, что как в митотических хромосомах, так и в интерфазных ядрах выявляются фибриллы хроматина с диаметром 30 нм. Хроматиновые фибриллы такого диаметра были видны как на ультратонких срезах после фиксации глутаровым альдегидом, так и на препаратах выделенного хроматина и выделенных хромосом в растворах, содержащих хотя бы низкие концентрации двухвалентных катионов. Все это говорило о том, что нуклеосомные цепочки хроматина каким-то специфическим образом уложены так, что возникает не хаотическая агрегация нуклеосом, а правильная нитчатая структура с диаметром 30 нм. Относительно характера упаковки нуклеосом в составе 30 нм фибриллы хроматина существует, по крайней мере, две точки зрения. Одна из них защищает, т.н. соленоидный тип укладки нуклеосом. Согласно этой модели, нить плотно упакованных нуклеосом диаметром 10 нм образует в свою очередь спиральные витки с шагом спирали около 10 нм. На один виток такой суперспирали приходится 6 нуклеосом. В результате такой упаковки возникает фибрилла спирального типа с центральной полостью, которая иногда на негативно окрашенных препаратах бывает видна как узкий «канал» в центре фибриллы. При частичном разворачивании, декомпактизации такой фибриллы и нанесении ее на подложку хорошо видно «зигзагообразное» расположение нуклеосом вдоль фибриллы. Считается, что гистон H1 обеспечивает взаимодействие между соседними нуклеосомами, не только сближая и связывая их друг с другом, но и обеспечивая кооперативную связь нуклеосом так, что образуется довольно плотная спираль из 10 нм фибриллы. Удаление, даже частичное, гистона H1 вызывает переход 30 нм фибриллы в 10 нм фибриллу, а полное удаление его вызывает разворачивание последней в структуру типа «бусин-на-нити». Такой соленоидный тип упаковки ДНК приводит к плотности упаковки равной приблизительно 40 (т.е. на каждый мкм нити приходится 40 мкм ДНК). Эти представления получили подтверждение при анализе структуры хроматина с помощью дифракции рентгеновских лучей и нейтронов. Здесь необходимо отметить, что представление о соленоидном типе укладки получены из анализа вторично конденсированного хроматина. Вначале были получены препараты хроматина в присутствии ЭДТА или выделялись в растворах низкой ионной силы в присутствии ионов магния. Во всех этих случаях первоначально хроматин деконденсировался до уровня «бусин на нити», где отсутствует или дестабилизируется контакт между нуклеосомами. Если же исследовать хроматин в составе ядер или в виде выделенных препаратов, но при поддержании определенной концентрации двухвалентных катионов (не ниже 1мМ), то можно видеть дискретность в составе 30 нм фибрилл хроматина: она состоит как бы из сближенных глобул того же размера, из нуклеомеров. В зарубежной литературе такие 30 нм глобулы или нуклеомеры получили название сверхбусин («супербиды»). Было обнаружено, что если в условиях, когда нуклеомерная структура фибрилл хроматина сохраняется, препараты хроматина подвергнуть нуклеазной обработке, то часть хроматина растворяется. При этом в раствор выходят частицы, имеющие размер около 30 нм с коэффициентом седиментации равным 45S в растворах, содержащих 1 мМ магния. Если такие выделенные нуклеомеры обработать ЭДТА, удалить ионы магния, то они разворачиваются в нуклеосомные цепочки, содержащие 6-8 нуклеосом. Таким образом, в состав одного нуклеомера входит отрезок ДНК, соответствующий 1600 парам оснований или 8 нуклеосомам. Компактность нуклеомера зависит от концентрации ионов магния и наличия гистона. Негистоновые белки в конформационных превращениях нуклеомеров не участвуют. Таким образом основная 30 нм фибрилла хроматина представляет собой линейное чередование нуклеомеров вдоль компактизованной молекулы ДНК. Вероятно, что гистоны H1, находясь в центральной зоне этой крупной частицы, взаимодействуя друг с другом, поддерживают ее целостность. В пользу этого говорят данные о кооперативном связывании гистонов H1 в группе по 6-8 молекул. Противоречие между соленоидной и нуклеомерной моделью упаковки нуклеосом в составе фибрилл хроматина может быть снято, если принять модель нерегулярного соленоида: число нуклеосом на виток спирали не является строго постоянной величиной, что может привести к чередованию участков с большим или меньшим числом нуклеосом на виток. Нуклеомерный уровень укладки хроматина обеспечивает 40 кратное уплотнение ДНК, что важно не только для достижения целей компактизации гигантских молекул ДНК. Компактизация ДНК в составе 30 нм фибрилл хроматина может налагать дополнительные функциональные ограничения. Так было обнаружено, что в составе 30 нм фибриллы хроматина ДНК становится практически недоступной для взаимодействия с таким ферментом как метилаза ДНК. Кроме того резко падает способность хроматина связываться с РНК-полимеразой и рядом регуляторных белков. Таким образом второй уровень компактизации ДНК может играть роль фактора, инактивирующего гены.В заключении необходимо еще раз напомнить, что как нуклеосомный, так и нуклеомерный (супербидный) уровни компактизации ДНК хроматина осуществляются за счет гистоновых белков, которые участвуют не только в образовании нуклеосом, но и в их кооперативном объединении в виде фибрилл ДНП, где ДНК претерпевает дополнительную сверхспирализацию. Все остальные уровни компактизации связаны с дальнейшим характером укладки 30 нм фибрилл в новые компактизационные уровни, где ведущую роль играют негистоновые белки. Врпрос 61. Структура ДНК. Модель Уотсона и Крика. | ||||||||||||||||||||||||||||