Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

Размножение делениемДеление свойственно прежде всего одноклеточным организмам. Как правило, оно осуществляется путём простого деления клетки надвое. У некоторых простейших (например, фораминифер) происходит деление на большее число клеток. Во всех случаях образующиеся клетки полностью идентичны исходной. Крайняя простота этого способа размножения, связанная с относительной простотой организации одноклеточных организмов, позволяет размножаться очень быстро. Так, в благоприятных условиях количество бактерий может удваиваться каждые 30—60 минут. Размножающийся бесполым путём организм способен бесконечно воспроизводить себя, пока не произойдёт спонтанное изменение генетического материала — мутация. Если эта мутация благоприятна, она сохранится в потомстве мутировавшей клетки, которое будет представлять собой новый клеточный клон. Размножение спорамиНередко бесполому размножению бактерий предшествует образование спор. Бактериальные споры — это покоящиеся клетки со сниженным метаболизмом, окружённые многослойной оболочкой, устойчивые к высыханию и другим неблагоприятным условиям, вызывающим гибель обычных клеток. Спорообразование служит как для переживания таких условий, так и для расселения бактерий: попав в подходящую среду, спора прорастает, превращаясь в вегетативную (делящуюся) клетку. Бесполое размножение с помощью одноклеточных спор свойственно и различным грибам и водорослям. Споры во многих случаях образуются путём митоза (митоспоры), причём иногда (особенно у грибов) в огромных количествах; при прорастании они воспроизводят материнский организм. Некоторые грибы, например злостный вредитель растений фитофтора, образуют подвижные, снабжённые жгутиками споры, называемые зооспорами или бродяжками. Проплавав в капельках влаги некоторое время, такая бродяжка «успокаивается», теряет жгутики, покрывается плотной оболочкой и затем, в благоприятных условиях, прорастает. Помимо митоспор, у многих из указанных организмов, а также у всех высших растений формируются споры и иного рода, а именно мейоспоры, образующиеся путём мейоза. Они содержат гаплоидный набор хромосом и дают начало поколению, обычно не похожему на материнское и размножающемуся половым путём. Таким образом, образование мейоспор связано с чередованием поколений — бесполого (дающего споры) и полового. Вегетативное размножение. Другой вариант бесполого размножения осуществляется путём отделения от организма его части, состоящей из большего или меньшего числа клеток. Из них развивается взрослый организм. Примером может служить почкование у губок и кишечнополостных или размножение растений побегами, черенками, луковицами или клубнями. Такая форма бесполого размножения обычно называется вегетативным размножением. В своей основе оно аналогично процессу регенерации. Играет важную роль в практике растениеводства. Так, может случиться, что высеянное растение (например, яблоня) обладает некой удачной комбинацией признаков. У семян данного растения эта удачная комбинация почти наверняка будет нарушена, так как семена образуются в результате полового размножения, а оно связано с рекомбинацией генов. Поэтому при разведении яблонь обычно используют вегетативное размножение — отводками, черенками или прививками почек на другие деревья. Бесполое размножение, воспроизводящее идентичные исходному организму особи, не способствует появлению организмов с новыми вариантами признаков, а тем самым ограничивает возможность приспособления видов к новым для них условиям среды. Средством преодоления этой ограниченности стал переход к половому размножению. Почкование некоторым видам одноклеточных свойственна такая форма бесполого размножения, как почкование. В этом случае происходит митотическое деление ядра. Одно из образовавшихся ядер перемещается в формирующееся локальное выпячивание материнской клетки, а затем этот фрагмент отпочковывается. Дочерняя клетка существенно меньше материнской, и ей требуется некоторое время для роста и достраивания недостающих структур, после чего она приобретает вид , свойственный зрелому организму.Почкование- вид вегетативного размножения. Почкованием размножаются многие низшие грибы, например дрожжи и даже многоклеточные животные, например пресноводная гидра. При почковании дрожжей на клетке образуется утолщение, постепенно превращающиеся в полноценную дочернюю клетку дрожжей. На теле гидры несколько клеток начинают делиться, и постепенно на материнской особи вырастает маленькая гидра, у которой образуются рот со щупальцами и кишечная полость, связанная с кишечной полостью „матери“.Если материнская особь поймает добычу, то часть питательных веществ попадает и в маленькую гидру,и наоборот, дочерняя особь, охотясь, также делится пищей с материнской особью. вскоре маленькая гидра отделяется от материнского организма и обычно располагается рядом с ней.(но не всегда!) Деление телом. Некоторые организмы могут размножаться делением тела на несколько частей, причём из каждой части вырастает полноценный организм, во всём сходный с родительской особью (плоские и кольчатые черви, иглокожие). 87.Половое размножение. Регулярные и нерегулярные формы. Половое размножение — процесс у большинства эукариот, связанный с развитием новых организмов из половых клеток (у одноклеточных эукариот при конъюгации функции половых клеток выполняют половые ядра). Образование половых клеток, как правило, связано с прохождением мейоза на какой-либо стадии жизненного цикла организма. В большинстве случаев, половое размножение сопровождается слиянием половых клеток, или гамет, при этом восстанавливается удвоенный, относительно гамет, набор хромосом. В зависимости от систематического положения эукариотических организмов, половое размножение имеет свои особенности, но как правило, оно позволяет объединять генетический материал от двух родительских организмов и позволяет получить потомков с комбинацией свойств, отсутствующей у родительских форм. Эффективности комбинирования генетического материала у потомков, полученных в результате полового размножения способствуют:
Такая форма полового размножения как партеногенез, не предусматривает слияния гамет. Но так как организм развивается из половой клетки (ооцита), партеногенез все равно считается половым размножением. Во многих группах эукариот произошло вторичное исчезновение полового размножения, или же оно происходит очень редко. В частности, в отдел дейтеромицетов (грибы) объединяет обширную группу филогенетических аскомицетов и базидиомицетов, утративших половой процесс. До 1888 года предполагалось, что среди наземных высших растений половое размножение полностью утрачено у сахарного тростника. Утеря полового размножения в какой-либо группе многоклеточных животных не описана. Однако известны многие виды (низшие ракообразные — дафнии, некоторые типы червей), способные в благоприятных условиях размножаться партеногенетически в течение десятков и сотен поколений. Например, некоторые виды коловраток на протяжении миллионов лет размножаются только партеногенетически, даже образуя при этом новые виды (!). У ряда полиплиодных организмов с нечётным числом наборов хромосом половое размножение играет малую роль в поддержании генетической изменчивости в популяции в связи с образованием несбалансированых наборов хромосом в гаметах и у потомков. Возможность комбинировать генетический материал при половом размножении имеет большое значение для селекции модельных и хозяйственно важных организмов. 88.Цитологические основы полового размножения. Мейоз, как специфический процесс при формировании половых клеток. Мейоз (от греч. meiosis — уменьшение) или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированных стволовых. С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации). Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений). 89.Гаметогенез. Строение половых клеток. Гаметогенез подразделяется на сперматогенез (процесс образования сперматозоидов у самцов) и оогенез (процесс образования яйцеклетки). По тому, что происходит с ДНК, эти процессы практически не отличаются: одна исходная диплоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с цитоплазмой, эти процессы кардинально различаются. В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем для развития зародыша, поэтому яйцеклетка – это очень крупная клетка, и когда она делится, цель – сохранить питательные вещества для будущего зародыша, поэтому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы цитоплазмы и при этом избавиться от ненужного генетического материала, от цитоплазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы, но позволяют поделить хромосомный набор. Полярные тельца отделяются при первом и втором делении мейоза 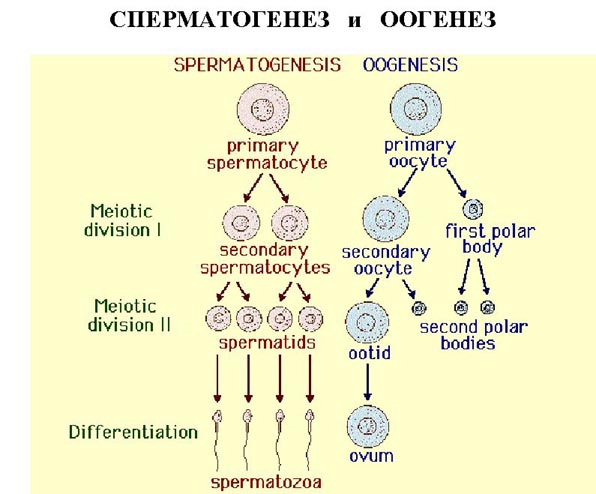 90.Закономерности сперматогенеза у млекопитающих и человека. Семенник состоит из нескольких тысяч цилиндрических семенных канальцев, в каждом из которых развиваются миллионы сперматозоидов. Стенки этих канальцев выстланы первичными, неспециализированными половыми клетками — сперматогониями. В течение эмбрионального развития и в детском возрасте сперматогонии делятся митотически, давая начало дополнительным сперматогониям, что ведет к росту семенника. После наступления половой зрелости некоторые из сперматогониев вступают в сперматогенез, состоящий из серии превращений, ведущих к образованию зрелых сперматозоидов. Другие сперматогонии продолжают делиться митотически и производят новые сперматогонии для последующих сперматогенезов. У большинства диких животных имеется определенный период, приходящийся на весну или на осень, когда семенники увеличиваются в размерах и в них происходит сперматогенез. В промежутке же между этими периодами семенник имеет небольшие размеры и содержит только сперматогонии. У человека и у большинства домашних животных сперматогенез после достижения половой зрелости протекает круглый год. Сперматогенез начинается с того, что сперматогонии вырастают и превращаются в более крупные клетки, известные под названием сперматоцитов первого порядка. Последние приступают к первому мейотическому делению, дающему начало двум одинаковым по величине клеткам — сперматоцитам второго порядка. В них происходит второе мейотическое деление, в результате которого образуются четыре одинаковые сперматиды. Сперматида — сферическая клетка, содержащая значительное количество цитоплазмы, — представляет собой зрелую гамету, так как она имеет гаплоидное число хромосом, но для того, чтобы превратиться в функционирующий сперматозоид, она должна приобрести обтекаемую форму, которая дает ей возможность плавать и встретиться с яйцом. Это связано со сложными процессами роста и видоизменения (не сопровождающимися, однако, клеточным делением). Ядро уменьшается в размерах и превращается в головку сперматозоида, который сбрасывает большую часть своей цитоплазмы. Некоторые из телец Гольджи собираются у переднего конца сперматозоида, образуя так называемую акросому, которая, возможно, помогает сперматозоиду прокалывать оболочку яйцеклетки. Обе имеющиеся в сперматиде центриоли перемещаются и располагаются непосредственно позади ядра. В ядре появляется небольшое углубление, и одна из центриолей — проксимальная — располагается в нем под прямым углом к оси сперматозоида. Вторая — дистальная — центриоль, лежащая позади проксимальной, дает начало осевой нити хвоста. Подобно осевым нитям жгутиков, эта нить состоит из двух продольных волокон, расположенных в середине, и окружающего их кольца из 9 пар продольных волокон. Митохондрии передвигаются к месту соединения головки и хвоста и образуют небольшой промежуточный участок (шейку), доставляющий энергию для биения хвостика. Большая часть цитоплазмы сперматиды отбрасывается; остается лишь тонкий слой, окружающий митохондрии в средней части и осевую нить хвостика. Сперматозоиды разных видов животных сильно различаются между собой. Почти все они имеют хвостик, но его величина и форма, так же как и особенности строения головки и шейки, могут быть весьма разнообразны. У некоторых животных, например у паразитического круглого червя аскариды, сперматозоиды не имеют хвостов и перемещаются при помощи амебоидных движений. У крабов и раков сперматозоиды необычные — у них нет хвоста, но на головке имеется три остроконечных выроста, которые прикрепляются к поверхности яйца и прочно удерживают сперматозоид. Шейка такого сперматозоида раскручивается подобно пружине и проталкивает его ядро в цитоплазму яйца, осуществляя таким образом оплодотворение. 91.Закономерности овогенеза у млекопитающих и человека. Первая фаза раневого процесса — фаза воспаления — характеризуется травматическим отеком тканей, повышением сосудистой проницаемости, ацидозом, миграцией лейкоцитов, тучных клеток и макрофагов. Происходит очищение раны путем фагоцитоза и лизиса некротизированных тканей. Во второй фазе раневого процесса — фазе регенерации — происходит развитие грануляционной ткани, постепенно заполняющей раневой дефект. Основными структурами этой ткани являются фибробласты, межклеточное вещество и капилляры. Клетки фиброблас-тического дифферона грануляционной ткани отличаются от фибробластов нормальной соединительной ткани высокой функциональной активностью. Они синтезируют белки и гликозаминоглнканы, образуя коллагеновые волокна. В развитии и созревании грануляционной ткани важную роль играют также макрофаги, тучные и плазматические клетки. Грануляционная ткань впоследствии преобразуется в рубцовую соединительную ткань. Третья фаза раневого процесса — фаза реорганизации рубца — характеризуется прогрессирующим уменьшением числа кровеносных сосудов и клеточных элементов (фибробластов, макрофагов, тучных клеток) при явлениях нарастания общей массы коллагеновых волокон. Параллельно с созреванием грануляционной ткани и ее превращением в рубцовую ткань происходит эпителизация раны. Эпителизация раны и созревание грануляционной ткани строго соответствуют во времени. В зависимости от характера и величины ранения, особенностей реактивности организма и других условий раневой процесс протекает по-разному. При небольшом объеме поражения имеет место заживление раны первичным натяжением. Воспаление и замещение дефекта в тканях следуют непосредственно за их травматическим отеком и не сопровождаются нагноением. К концу первой недели раневой процесс в основном завершается. Если же объем поражения велик и края раны оказываются на более или менее значительном расстоянии друг от друга, то заживление раны осуществляется через нагноение с образованием хорошо развитой грануляционной ткани с последующим ее рубцеванием. Заживление раны происходит вторичным натяжением со значительной продолжительностью фаз раневого процесса. Заживление раны первичным и вторичным натяжением имеет количественные, но не качественные различия. Механизмы регенерации при этом принципиально сходны и включают воспаление, разрастание соединительной ткани, эпителизацию. Знание ключевых звеньев регенерацbонного процесса позволяет целенаправленно вести поиск средств регуляции заживления ран и разработке способов тканевой терапии. |