Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

Условная классификация модификационной изменчивости
Механизм модификационной изменчивостиОкружающая среда как причина модификацийМодификационная изменчивость — это результат не изменений генотипа, а его реакции на условия окружающей среды. При модификационной изменчивости наследственный материал не изменяется, — изменяется проявление генов. Под действием определенных условий окружающей среды на организм изменяется течение ферментативных реакций (активность ферментов) и может происходить синтез специализированных ферментов, некоторые из которых (MAP-киназа и др.) ответственны за регуляцию транскрипции генов, зависящую от изменений окружающей среды. Таким образом, факторы окружающей среды способны регулировать экспрессию генов, то есть интенсивность выработки ими специфических белков, функции которых отвечают специфическим факторам окружающей среды. Например, за выработку меланина ответственны четыре гена, которые находятся в разных хромосомах. Наибольшее количество доминантных аллелей этих генов — 8 — содержится у людей негроидной расы. При воздействии специфической окружающей среды, например, интенсивного воздействия ультрафиолетовых лучей, происходит разрушение клеток эпидермиса, что приводит к выделению эндотелина-1 и эйкозаноидов. Они вызывают активацию фермента тирозиназы и его биосинтез. Тирозиназа, в свою очередь, катализирует окисление аминокислоты тирозина. Дальнейшее образование меланина проходит без участия ферментов, однако большее количество фермента обуславливает более интенсивную пигментацию. Норма реакции: предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет предел для каждого вида — например, усиленное кормление приведет к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и наследуется. Для разных изменений есть разные пределы нормы реакции. Например, сильно варьируют величина удоя, продуктивность злаков (количественные изменения), слабо — интенсивность окраски животных и т. д. (качественные изменения). В соответствии с этим норма реакции может быть широкой (количественные изменения — размеры листьев многих растений, размеры тела многих насекомых в зависимости от условий питания их личинок) и узкой (качественные изменения — окраска у куколок и имаго некоторых бабочек). Тем не менее, для некоторых количественных признаков характерна узкая норма реакции (жирность молока, число пальцев на ногах у морских свинок), а для некоторых качественных признаков — широкая (например, сезонные изменения окраски у многих видов животных северных широт). Характеристика модификационной изменчивости
67.Рекомбинация наследственного материала в генотипе. Комбинативная изменчивость. Мейоз и оплодотворение обеспечивают получение организмами нового поколения эволюционно сложившегося, сбалансированного по дозам генов наследственного материала, на основе которого осуществляется развитие организма и отдельных его клеток. Благодаря этим двум механизмам в ряду поколений особей данного вида формируются определенные видовые характеристики и вид как реальная единица живой природы существует продолжительное время. Однако у разных представителей вида в силу постоянно идущего мутационного процесса один и тот же набор генов генома представлен разными их аллелями. Так как при половом размножении у многих видов в воспроизведении потомства принимают участие две особи, то совершенно очевидно, что в результате оплодотворения разные зиготы получают неодинаковый набор аллелей в их генотипах. Увеличению генотипического разнообразия представителей вида способствуют также механизмы, приводящие к перекомбинации родительских аллелей особи в ее гаметах. Действительно, если бы гаметы, образуемые организмом, были одинаковы по набору аллелей в их геноме, то у потомков одной пары организмов при раздельнополости или одного гермафродитного организма не наблюдалось бы генотипического разнообразия. В каждом новом поколении вида генотипически различными были бы лишь дети разных родителей. Реально в природе наблюдается разнообразие потомков одних и тех же родителей. Например, родные братья и сестры различаются не только по полу, но и по другим признакам. Такие различия потомков объясняются тем, что в каждом акте оплодотворения встречаются генетически различающиеся гаметы. Механизмом,обеспечивающим разнообразие гамет, образуемых одним и тем же организмом, является мейоз, в ходе которого происходит не только уменьшение вдвое наследственного материала, попадающего в гаметы, но и эффективное перераспределение родительских аллелей между гаметами. Процессами, приводящими к перекомбинации генов и целых хромосом в половых клетках, являются Кроссинговер и расхождение бивалентов в анафазе I мейоза (см. гл. 5). Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом (рис. 3.72). Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах. 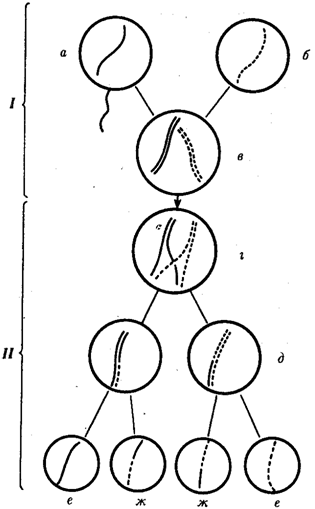 Рис. 3.72. Кроссинговер как источник генетического разнообразия гамет: I — оплодотворение родительских гамет а и б с образованием зиготы в; II — гаметогенез в организме, развившемся из зиготы в; г — кроссинговер, происходящий между гомологами в профазе I; д — клетки, образовавшиеся после 1-го мейотического деления; е, ж — клетки, образовавшиеся после 2-го деления мейоза (е — некроссоверные гаметы с исходными родительскими хромосомами; ж — кроссоверные гаметы с перекомбинацией наследственного материала в гомологичных хромосомах) Понятно, что кроссинговер как механизм рекомбинации эффективен лишь в том случае, когда соответствующие гены отцовской и материнской хромосом представлены разными аллелями. Абсолютно идентичные группы сцепления при кроссинговере не дают новых сочетаний аллелей. Кроссинговер происходит не только в предшественницах половых клеток при мейозе. Он наблюдается также в соматических клетках при митозе. Соматический кроссинговер описан у дрозофилы, у некоторых видов плесеней. Он осуществляется в ходе митоза между гомологичными хромосомами, однако его частота в 10 000 раз меньше частоты мейотического кроссинговера, от механизма которого он ничем не отличается. В результате митотического кроссинговера появляются клоны соматических клеток, различающихся по содержанию в них аллелей отдельных генов. Если в генотипе зиготы данный ген представлен двумя разными аллелями, то в результате соматического кроссинговера могут появиться клетки с одинаковыми либо отцовскими, либо материнскими аллелями данного гена (рис. 3.73). 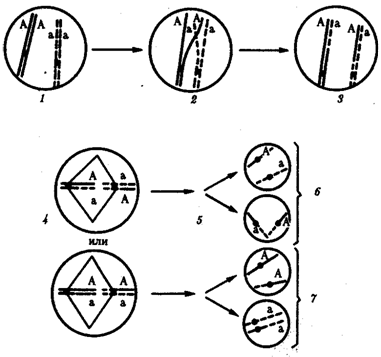 Рис. 3.73. Кроссинговер в соматических клетках: 1 — соматическая клетка, в гомологичных хромосомах которой ген А представлен двумя разными аллелями (А и а); 2 — кроссинговер; 3 — результат обмена соответствующими участками между гомологичяыми хромосомами; 4 — расположение гомологов в плоскости экватора веретена деления в метафазе митоза (два варианта); 5 — образование дочерних клеток; 6 — образование гетерозитотиых по гену А клеток, сходных с материнской клеткой по набору аллелей (Аа); 7 — образование гомозиготных по гену А клеток, отличающихся от материнской клетки по набору аллелей (АА или аа) Расхождение бивалентов в анафазе I мейоза. В метафазе I мейоза в экваториальной плоскости ахромативнового веретена выстраиваются биваленты, состоящие из одной отцовской и одной материнской хромосомы. Расхождение гомологов, которые несут разный набор аллелей генов в анафазе I мейоза, приводит к образованию гамет, отличающихся по аллельному составу отдельных групп сцепления (рис. 3.74). 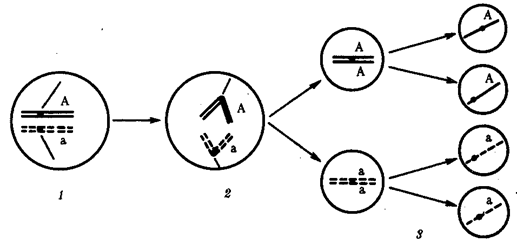 Рис. 3.74. Расхождение гомологичных хромосом в анафазе I мейоза как источник генетического разнообразия гамет: 1—метафаза I мейоза (расположение бивалента в плоскости экватора веретена деления); 2 — анафаза I мейоза (расхождение гомологов, несущих разные аллели гена А к разным полюсам); 3 — второе мейотическое деление (образование двух типов гамет, различающихся по аллелям гена А) 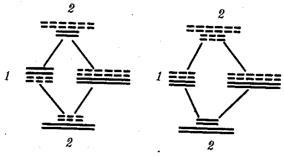 Рис. 3.75. Случайный характер расположения бивалентов в метафазе (1) и независимое расхождение их в анафазе (2) первого мейотического деления В связи с тем что ориентация бивалентов по отношению к полюсам веретена в метафазе I оказывается случайной, в анафазе I мейоза в каждом отдельном случае к разным полюсам направляется гаплоидный набор хромосом, содержащий оригинальную комбинацию родительских групп сцепления (рис. 3.75). Разнообразие гамет, обусловленное независимым поведением бивалентов, тем больше, чем больше групп сцепления в геноме данного вида. Оно может быть выражено формулой 2n, где п — число хромосом в гаплоидном наборе. Так, у дрозофилы п = 4 и количество типов гамет, обеспечиваемое перекомбинацией родительских хромосом в них, равно 24 = 16. У человека п = 23, и разнообразие гамет, обусловленное этим механизмом, соответствует 223, или 8388608. Кроссинговер и процесс расхождения бивалентов в анафазе I мейоза обеспечивают эффективную рекомбинацию аллелей и групп сцепления генов в гаметах, образуемых одним организмом. Оплодотворение. Случайная встреча разных гамет при оплодотворении приводит к тому, что среди особей вида практически невозможно появление двух генотипически одинаковых организмов. Достигаемое с помощью описанных процессов генотипическое разнообразие особей предполагает наследственные различия между ними на базе общего видового генома. Таким образом, геном как высший уровень организации наследственного материала благодаря мейозу и оплодотворению сохраняет свои видовые характеристики. Но одновременно эти же процессы обеспечивают индивидуальные наследственные различия особей, в основе которых лежит рекомбинация генов и хромосом, т.е. комбинативную изменчивость. Комбинативная изменчивость, проявляющаяся в генотипическом разнообразии особей, повышает выживаемость вида в изменяющихся условиях его существования. 68.Мутационная изменчивость и её виды. Мутацией (от лат. mutatio - перемена) называется изменение, обусловленное реорганизацией воспроизводящих структур клетки, изменением ее генетического аппарата. Этим мутации резко отличаются от модификаций, не затрагивающих генотип . особи. Мутации возникают внезапно, скачкообразно и иногда резко отличают организм от исходной формы. Растениеводам и животноводам такие изменения были известны давно. Ряд наследственных изменений описал Дарвин в труде «Изменчивость животных и растений в условиях одомашнения» (1868). Мутационной изменчивости посвятили свои работы С. И. Коржинский (1899) в де Фриз (1901), последнему из них принадлежит термин «мутации». В настоящее время известны мутации у всех классов животных растений и вирусов (рис. 56, 57). Существует много мутаций и у человека. Мутационная изменчивость свойственна всем организмам, она поставляет материал для отбора, с ней связана эволюция - процесс образования новых видов, сортов и пород. По характеру изменений генетического аппарата различают мутации, обусловленные: 1) изменением числа хромосом (полиплоидия, гетероплоидия, гаплоидия); 2) изменением структуры хромосом (хромосомные абберации); 3) изменением молекулярном структуры гена (генные или точковые мутации). 69.Соматические мутации. Понятие о клеточных клонах. Понятие о мозаицизме. Соматические мутации, мутации, возникающие в клетках тела и обусловливающие мозаичность организма, т. е. образование в нём отдельных участков тела, тканей или клеток с отличным от остальных набором хромосом или генов. В клетках развивающегося организма могут возникать Соматические мутации всех тех типов, которые наблюдаются в половых клетках: умножения хромосомного набора в целом в результате нормального деления хромосом без последующего деления ядра и клетки (см. Полиплоидия); трисомии и моносомии различных хромосом в результате отхождения двух дочерних хромосом к одному полюсу (вместо расхождения их к разным полюсам); потери хромосомы в одной из дочерних клеток в результате её задержки в зоне экваториальной пластинки при делении и т.д. В соматических клетках с той или иной частотой имеют место инверсии (перевороты), делеции (утраты) и транслокации (перестановки) участков хромосом, а также мутации отдельных генов. Чем раньше в процессе развития организма возникает Соматические мутации, тем большее количество клеток-потомков её унаследует при условии, что мутация не убивает клетку-носительницу и не снижает темпов её размножения. Генные Соматические мутации проявляются относительно редко, т.к. в подавляющем большинстве случаев функция мутантного гена или выпавшего участка хромосомы компенсируется наличием нормального гомологичного гена или нормального участка в партнёре - гомологе мутантной хромосомы. Проявление некоторых Соматические мутации подавляется соседством нормальной ткани. Наконец, Соматические мутации может не проявиться в силу того, что в данной ткани соответствующий участок хромосомы неактивен. Тем не менее в начале 60-х гг. 20 в. выяснилось важное значение Соматические мутации в патогенезе ненормального развития половой системы, в возникновении самопроизвольных абортов и врождённых уродств, в канцерогенезе. Мозаицизм (генетический мозаицизм, хромосомный мозаицизм — mosaicism; мозаичность; могут употребляться синонимы «мозаичная форма», «мозаичный кариотип») — от фр. mosaique «мозаика» — наличие в тканях (растения, животного, человека) генетически различающихся клеток. Следует отличать мозаицизм от химеризма, при котором два (или более) генотипа происходят более чем от одной зиготы. Понятие мозаицизма связано с понятиями трисомии и анеуплоидии. Может возникать в результате:
70.Генеративные мутации. Изменения наследственной программы половых клеток человека приводят к рождению потомства с различными наследственно обусловленными болезнями, в зависимости от ранга мутаций — генными или хромосомными. Различные генные мутации по-разному сказываются на жизнеспособности организма, причем в случае их рецессивности они могут долго не проявляться фенотипически у потомков. Хромосомные перестройки и геномные мутации приводят к выраженным отклонениям в развитии и часто являются причиной гибели организма на разных стадиях его онтогенеза, обычно в раннем эмбриогенезе. В значительной степени именно этими мутациями определяется высокий процент (15%) прерывания диагностированных беременностей. Триплоидии плода, как правило, приводят к прерыванию беременности на ранних стадиях, однако описано очень небольшое число случаев живорождения триплоидов. Анэуплоидия по разным хромосомам встречается как в материале абортусов, так и у рожденных детей. Некоторые анэуплоидий несовместимы с жизнью. Так, трисомия по 16-й хромосоме обнаруживается только в материале абортусов. В то же время у человека известны синдромы, связанные с аномалиями числа хромосом, характеризующиеся разной степенью жизнеспособности. Наиболее частым хромосомным заболеванием у человека является синдром Дауна, обусловленный три-сомией по 21-й хромосоме, встречающийся с частотой 1—2 на 1000 (рис. 4.3). Примерно в 60% случаев трисомия 21 является причиной гибели плода, около 30% родившихся умирает на первом году жизни. Еще 46% не переживает Злетний рубеж, однако иногда люди с синдромом Дауна доживают до значительного возраста (рис. 4.4), хотя в целом продолжительность их жизни сокращена. Применение эффективных противомикробных препаратов позволяет несколько увеличить продолжительность жизни таких больных. Трисомия 21 может быть результатом случайного нерасхождения гомологичных хромосом в мейозе. Наряду с этим известны случаи регулярной трисомии, связанной с транслокацией 21-й хромосомы на другую —21, 22, 13, 14 или 15-ю хромосому (рис. 4.5). Среди других аутосомных трисомий известны трисомии по 13-й хромосоме — Синдром Патау (рис. 4.6), а также по 18-й хромосоме — синдром Эдвардса (рис. 4.7), при которых жизнеспособность новорожденных резко снижена. Они гибнут в первые месяцы жизни из-за множественных пороков развития. Применение методов дифференциального окрашивания хромосом позволило открыть три новых синдрома, обусловленных трисомиями по 8, 9 и 22-й хромосомам, при которых также наблюдаются тяжелые комплексные пороки развития (рис. 4.8). Достаточно часто у человека встречаются анэуплоидии по половым хромосомам (рис. 4.9—4.11). В отличие от анэуплоидии по аутосомам дефекты умственного развития у больных выражены не столь отчетливо, у многих оно в пределах нормы, а иногда даже выше среднего. Вместе с тем у них постоянно наблюдаются нарушения развития половых органов и гормонозависимого роста тела. Реже встречаются пороки развития других систем. Относительно благоприятные последствия увеличения числа Х-хромосом, видимо, связаны с возможностью компенсации дозы соответствующих генов благодаря естественной генетической инактивации этих хромосом, а также мозаичному характеру такой инактивации. Среди анэуплоидных синдромов по половым хромосомам моносомия Х (ХО) (синдром Шерешевского — Тернера) встречается много реже, чем трисомия X, синдром Клайнфельтера (XXY, XXXY), а также XYY, что указывает на наличие сильного отбора против гамет, не содержащих половых хромосом, или против зигот ХО. Это предположение подтверждается достаточно часто наблюдаемой моносомией Х среди спонтанно абортированных зародышей. В связи с этим допускается, что выжившие зиготы ХО являются результатом не мейотического, а митотического нерасхождения, или утраты Х-хромосомы на ранних стадиях развития (см. рис. 4.9). Моносомии YO у человека не обнаружено. Из синдромов, связанных со структурными аномалиями хромосом, известен транслокационный синдром Дауна (см. рис. 4.5), при котором число хромосом в кариотипе формально не изменено и равно 46, так как дополнительная 21-я хромосома транслоцирована на одну из акроцентрических хромосом. При транслокации длинного плеча 22-й хромосомы на 9-ю развивается хронический миелолейкоз. При делении короткого плеча 5-й хромосомы развивается синдром кошачьего крика, при котором наблюдаются общее отставание в развитии, низкая масса при рождении, лунообразное лицо с широко расставленными глазами и характерный плач ребенка, напоминающий кошачье мяукание, причиной которого является недоразвитие гортани (рис. 4.12). 71.Виды мутаций. Спонтанные и индуцированные. Классификация мутагенов. 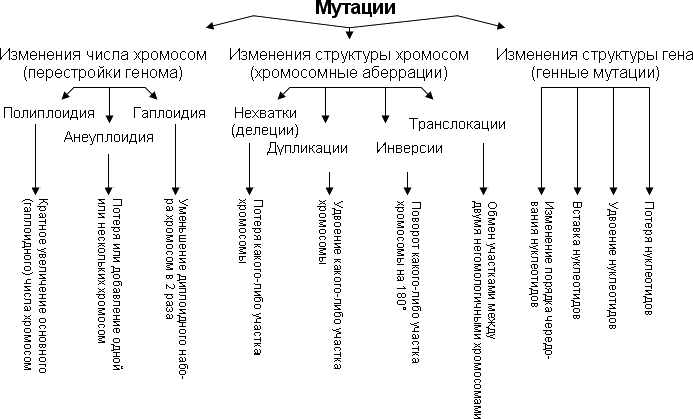 72.Геномные мутации. Болезни, связанные с нарушением количества аутосом. Геномные мутации связаны с изменением числа хромосом. Например, у растений довольно часто обнаруживается явление полиплоидии - кратного изменения числа хромосом. У полиплоидных организмов гаплоидный набор хромосом n в клетках повторяется не 2, как у диплоидов, а значительно большее число раз (3n, 4п, 5п и до 12n). Полиплоидия - следствие нарушения хода митоза или мейоза: при разрушении веретена деления удвоившиеся хромосомы не расходятся, а остаются внутри неразделившейся клетки. В результате возникают гаметы с числом хромосом 2n. При слиянии такой гаметы с нормальной (n) потомок будет иметь тройной набор хромосом. Если геномная мутация происходит не в половых, а в соматических клетках, то в организме возникают клоны (линии) полиплоидных клеток. Нередко темпы деления этих клеток опережают темпы деления нормальных диплоидных клеток (2n). В этом случае быстро делящаяся линия полиплоидных клеток образует злокачественную опухоль. Если она не будет удалена или разрушена, то за счет быстрого деления полиплоидные клетки вытеснят нормальные. Так развиваются многие формы рака. Разрушение митотического веретена может быть вызвано радиацией, действием ряда химических веществ - мутагенов . Геномные мутации в животном и растительном мире многообразны, но у человека обнаружены только 3 типа геномных мутаций: тетраплоидия , триплоидия и анеуплоидия . При этом из всех вариантов анеуплоидий встречаются только трисомии по аутосомам , полисомии по половым хромосомам (три-, тетра- и пентасомии), а из моносомий встречаются только моносомия-Х. |