Содержание 14. А
 Скачать 3.72 Mb. Скачать 3.72 Mb.
|

|
Активный транспорт, натриевый насос. В предыдущем разделе описаны пассивная диффузия ионов и образующийся вследствие этого мембранный потенциал при заданных внутри- и внеклеточных концентрациях ионов. Однако в результате этого процесса концентрация ионов внутри клетки стабилизируется не автоматически, поскольку мембранный потенциал несколько более электроотрицателен, чем ЕK, и намного - по сравнению с ENa (около +60 мВ). Благодаря диффузии внутриклеточные концентрации ионов, по крайней мере калия и натрия, должны уравниваться с внеклеточными. Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая-калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии. 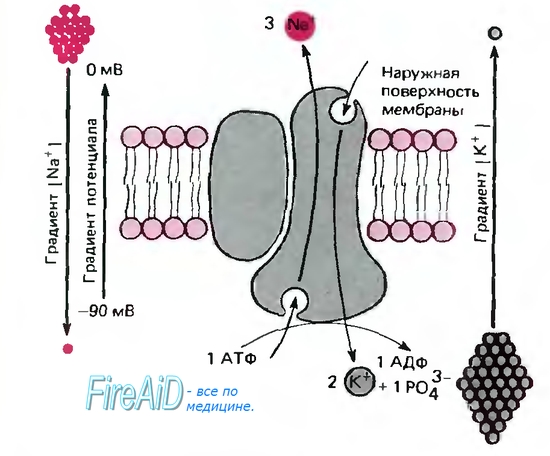 Шпора 8. Вопрос 15 . Поверхностный аппарат клетки. Транспорт макромолекул. Пузырьки осуществляют транспорт различных веществ путем быстрого и непрерывного челночного перемещения между различными окруженными мембраной органеллами эукариотической клетки. Экзоцитоз и эндоцитоз через плазматическую мембрану - лишь наиболее известные примеры этого сложного процесса. Несмотря на важность везикулярного транспорта , довольно мало известно о молекулярных механизмах, приводящих в действие, направляющих и регулирующих его. Очевидно, что для подобных процессов требуется энергия, возможно, в виде гидролиза АТР. Известно, что движение пузырьков в цитоплазме направляется микротрубочками и осуществляется белками-транслокаторами. На цитоплазматических поверхностях пузырьков имеются, очевидно, специальные молекулы- маркеры , направляющие пузырьки к соответствующей мембране, однако о природе и разнообразии этих молекул ничего не известно. Можно предположить, что непрерывное слияние мембран , имеющее место при опосредуемом пузырьками транспорте, приводит к значительному перемешиванию мембранных компонентов, однако различные мембраны в клетке сохраняют свой характерный состав. Подобное постоянство означает, что в клетке должны существовать специальные механизмы, восстанавливающие мембраны после их перемешивания . Вопрос №16. Рецепторная роль плазмолеммы. Рецептор-молекулы(гликопротеиды)способные распознавать и избирательно связывать химические в-ва лиганды. В роли лигандов могут быть питательные вещества, и вещества необходимые для других метаболических процессов (сборка структур) ЛНП источник холестерина, чужеродные в-ва (антигены), которые подлежат уничтожению, сигнальные молекулы (гормоны, нейромедиаторы). Сигнальные молекулы могут быть: гидрофильные и гидрофобные. Гидрофобные (стероидные гормоны)- стероиды проникают через мембрану клетки связываются в внутриклеточными рецепторами и действуют прямо на ДНК. Гидрофильные (нейромедиаторы) это гормоны белковой природы, такие как инсулини ряд других (адренолин). Связываются с рецепторами клеточной мембраны. Вопрос №17 Особенности воздействия на клетки-мишени гидрофильных и гидрофобных сигнальных молекул. Клетки-мишени- это клетки, которые специфически взаимодействуют с гормонами с помощью специальных белков-рецепторов. Эти белки-рецепторы располагаются на наружной мембране клетки, или в цитоплазме, или на ядерной мембране и на других органеллах клетки. Любой белок-рецептор состоит, минимум из двух доменов (участков), которые обеспечивают выполнение двух функций: узнавание гормона; преобразование и передачу полученного сигнала в клетку. Большинство рецепторов изучены недостаточно, потому что их выделение и очистка очень сложные, а содержание каждого вида рецепторов в клетках очень низкое. Но известно, что гормоны взаимодействуют со своими рецепторами физико-химическим путем. Между молекулой гормона и рецептором формируются электростатические и гидрофобные взаимодействия. При связывании рецептора с гормоном происходят конформационные изменения белка-рецептора и комплекс сигнальной молекулы с белком-рецептором активируется. В активном состоянии он может вызывать специфические внутриклеточные реакции в ответ на принятый сигнал. Если нарушен синтез или способность белков-рецепторов связываться с сигнальными молекулами, возникают заболевания - эндокринные нарушения. Механизмы действия гормонов на клетки-мишени. В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране. Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет.Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору. Циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия: 1.аденилатциклазная (или гуанилатциклазная) системы; 2.фосфоинозитидный механизм. Аденилатциклазная система.Основные компоненты: мембранный белок-рецептор, G-белок, фермент аденилатциклаза, гуанозинтрифосфат, протеинкиназы, АТФ. Белок-рецептор, G-белок, рядом с которым располагаются ГТФ и фермент (аденилатциклаза) встроены в мембрану клетки. Комплекс "G-белок-ГТФ" активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ. Она обладает способностью активировать особые ферменты - протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ. При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного белка. В различных типах клеток фосфорилированию в результате активации аденилат-циклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты, ядерные белки, мембранные белки. В результате реакции фосфорилирования белки могут становятся функционально активными или неактивными.Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ. Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс "GTP-G-белок" ингибирует аденилатциклазу.Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме усиливается действие гормона. Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата. Вопрос №18. Сигнализация с участием рецепторов клеточной поверхности и вторичных посредников на примере с-АТФ. Шпора 21. Вопрос 19. Система сигнализации: эндокринная, синоптическая. Роль медиаторов и гормонов. Шпора 10. Роль медиаторов: обеспечение механической прочности, обмен вещест, транспорт веществ Вопрос 20. Понятие о вторичных посредниках. Инозитолфосфатная система Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С, Gрlс - белок, активирующий фосфолипазу С, белки и ферменты мембран и цитозоля. Последовательность событий, приводящих к активации фосфолипазы С: связывание сигнальной молекулы, например гормона с рецептором (R) вызывает изменение конформации и увеличение сродства к Gplc-белку. образование комплекса [Г] [R] [Gрlс ГДФ] приводит к снижению сродства α-протомера G рlс белка к ГДФ и увеличению сродства к ГТФ. ГДФ заменяется на ГТФ. это вызывает диссоциацию комплекса; отделившаяся α-субъединица, связанная с молекулой ГТФ, приобретает сродство к фосфолипазе С. α-ГТФ взаимодействует с фосфолипазой С и активирует её. Под действием фосфолипазы С происходит гидролиз липида мембраны фосфатидилинозитол-4,5 - биофосфата (ФИФ2). в ходе гидролиза образуется и выходит в цитозоль гидрофильное вещество инозитол-1,4,5-трифосфат (ИФ3). Другой продукт реакции диацилглицерол (ДАГ) остаётся в мембране и участвует в активации фермента протеинкиназы С (ПКС). инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2 - канала мембраны ЭР, это приводит к изменению конформации белка и открытию канала - Са²+ поступает в цитозоль. В отсутствие в цитозоле ИФ3 канал закрыт. Активация протеинкиназы С. • Повышение концентрации Са²+ в цитозоле клетки увеличивает скорость взаимодействия Са²+ с неактивным цитозольным ферментом протеинкиназой С(ПКС) и белком кальмодулином, таким образом сигнал, принятый рецептором клетки, раздваивается. • Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция. • На внутренней стороне мембраны образуется ферментативный комплекс - [ПКС] [Са²+] [ДАГ] [ФС] - активная протеинкиназа С, фосфорилирующая специфические ферменты по серину и треонину. Вопрос 21. Понятие о вторичных посредниках. Аденилатцеклазная система Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В нем задействовано мимимум пять хорошо изученных белков: 1) рецептор гормона; 2) фермент аденилатциклаза, выполняющая функцию синтеза циклического АМФ (цАМФ); 3) G-белок, осуществляющий связь между аденилатциклазой и рецептором; 4) цАМФ-зависимая протеинкиназа, катализирующая фосфорилирование внутриклеточных ферментов или белков-мишеней, соответственно изменяя их активность; 5) фосфодиэстераза, которая вызывает распад цАМФ и тем самым прекращает (обрывает) действие сигнала (рис. 8.5). Получены в чистом виде α- и β-адренергические рецепторы из плазматических мембран клеток печени, мышц и жировой ткани. Показано, что связывание гормона с β-адренергическим рецептором приводит к структурным изменениям внутриклеточного домена рецептора, что в свою очередь обеспечивает взаимодействие рецептора со вторым белком сигнального пути – ГТФ-связывающим. ГТФ-связывающий белок – G-белок – представляет собой смесь 2 типов белков: активного Gs(от англ. stimulatory G) и ингибиторного Gi с мол. массой 80000–90000. В составе каждого из них имеется три разные субъединицы (α-, β- и γ-), т.е. это гетеротримеры. Показано, что β-субъеди-ницы Gsи Giидентичны (мол. масса 35000); в то же время α-субъединицы, являющиеся продуктами разных генов (мол. масса 45000 и 41000), оказались ответственными за проявление G-белком активаторной и ингибиторной активности соответственно. Гормонрецепторный комплекс сообщает G-белку способность не только легко обменивать эндогенный связанный ГДФ на ГТФ, но и переводить Gs-белок в активированное состояние, при этом активный G-белок диссоциирует в присутствии ионов Mg2+на β-, γ-субъединицы и комплекс α-субъединицы Gsв ГТФ-форме; этот активный комплекс затем перемещается к молекуле аденилатциклазы и активирует ее. Сам комплекс затем подвергается самоинактивации за счет энергии распада ГТФ и реассоциации β- и γ-субъединиц с образованием первоначальной ГДФ-формы Gs. Вопрос №22 межклеточные контакты. Адгезивне соединения. Межклеточные точечные сцепляющие соединения обнаружены у многих неэпителиальных тканей, но более отчетливо описана структура специальных (адгезивных) лент в однослойных эпителиях. Это структура опоясывает весь периметр эпителиальной клетки, подобно тому как это происходит в случае плотного соединения. Чаще всего такой поясок или лента лежит ниже плотного соединения. В этом месте плазматические мембраны не сближены, а даже несколько раздвинуты на расстояние 25-30 нм, и между ними видна зона повышенной плотности. Это ничто иное как места взаимодействия трансмембранных гликопротеидов, которые специфически сцепляются друг с другом и обеспечивают механическое соединение мембран двух соседних клеток. Эти линкерные белки относятся к Е-кадгеринам - белкам, обеспечивающим специфическое узнавание клетками однородных мембран. Разрушение этого слоя гликопротеидов приводит к обособлению отдельных клеток и разрушению эпителиального пласта. С цитоплазматической стороны около мембраны видно скопление какого-то плотного вещества, к которому примыкает слой тонких (6-7 нм) филаментов, лежащих вдоль плазматической мембраны в виде пучка, идущего по всему периметру клетки. Тонкие филаменты относятся к актиновым фибриллам, они связываются с плазматической мембраной посредством белка катенина, образующего плотный около мембранный слой. Функциональное значение такого ленточного соединения заключается на только в механическом сцеплении клеток друг с другом: при сокращении актиновых филаментов в ленте может изменяться форма клетки. Считается, что кооперативное сокращение актиновых фибрилл во всех клетках эпителиального пласта может вызвать изменение его геометрии, например, сворачивание в трубку, подобно тому, что происходит при образовании нервной трубки у эмбрионов позвоночных. Шпора 10. Вопрос №23.межклеточные соединения. Проводящие (химические)контакты Нексусы (щелевидные контакты):
Синапсы — см. раздел по нервной ткани. Обеспечивают передачу потенциала действия (нервного импульса) с нервной клетки на другую нервную или иную клетку. 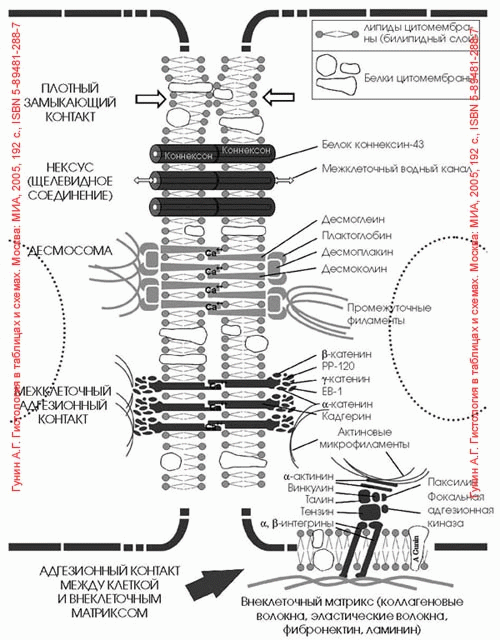 Шпора 10 Вопрос 24. Синаптическая передача нервного импульса. Синаптическая передача (также называемая нейропередача) — электрические движения в синапсах вызванные распространением нервных импульсов. Каждая нервная клетка получает медиатор из пресинаптического нейрона или из терминального окончания или из постсинаптичекского нейрона или дендрида вторичного нейрона и посылает его обратно нескольким нейронам, которые повторяют данный процесс, таким образом, распространяя волну импульсов до тех пор, пока импульс не достигнет опроделенного органа или специфической группы нейронов. Нервные импульсы необходимы для распространения сигналов. Эти сигналы посылаются в и исходят из центральной нервной системы через эфферентные и афферентные нейроны для координации гладких, скелетных и сердечных мышц, секреции желез и функционирования органов, важных для долгосрочного выживания многоклеточных позвоночных организмов, таких как млекопитающие. Нейроны образуют нейронные сети, по которым передаются нервные импульсы. Каждый нейрон образует не менее 15,000 соединений с другими нейронами. Нейроны не соприкасаются друг с другом; они образуют точки соприкосновения, называемые синапсами. Нейрон передают информацию с помощью нервного импульса. Когда импульс нейрона достигает синапса это приводит к выделению медиаторов, которые влияют на другие клетки, приводя к угнетению или облегчению. Следующий нейрон может соединяться с множеством других нейронов, и если возбуждающие процессы превалируют над угнетающими, то будет развит потенциал действия в основании аксона, таким образом передавая информацию к следующему нейрону, приводя к памяти или действию. Примером распространения с помощью нейронов является сердечное сокращение. Сокращение осуществляется когда сигнал поступает из синоатриального узла с частотой, заставляющей сердце полностью сократиться, выбросив всю кровь и последующему наполнению новой порцией крови. Важно, что что импульс опосылается из синоатриального узла, так как направление импульса между нейронами заставляет мышцу сокращаться полностью. Если импульс будет поступать из синоатриального узла сердце будет сокращаться неуверенно и не будет выбрасывать всю кровь в систему. Вопрос №25. Межклеточные соединения. Плотные или замыкающие контакты Шпора 10 В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания. Вопрос №26.метаболический аппарат клетки. Этапы внутриклеточного транспорта. МЕТАБОЛИЗМ - это обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода. Ассимиля́ция — совокупность процессов анаболизма (биосинтеза) в живом организме, в ходе которых различные вещества включаются в его состав. В ходе ассимиляции простые вещества (сложные первоначально расщепляются до простых), неспецифические для какого-либо организма, превращаются в сложные, характерные для данного вида соединения (усваиваются). Ассимиляция уравновешивается суммой процессов диссимиляции (распада). Метаболический аппарат клетки - совокупность ее структур, участвующих в метаболизме. Органоиды, участвующие в ассимиляции: шЭПС- На каналах шероховатой ЭПС в виде полисом расположены рибосомы. Здесь протекает синтез белков, преимущественно продуцируемых клеткой на экспорт (удаление ич клегки), например, секретов железистых клеток. Здесь же происходят образование липи-дов и белков цитоплазматической мембраны и их сборка. Плотно упакованные цистерны и каналы гранулярной ЭПС образуют слоистую структуру, где наиболее активно протекает синтез белка. Это место называется эргастоплазмой. Гладкая ЭПС-на мембранах гладкой ЭПС рибосом нет. Здесь протекает в основном синтез жиров и подобных им веществ (например, стероидных гормонов), а также углеводов. По каналам гладкой ЭПС также происходит перемещение готового материала к месту его упаковки в гранулы (в зону комплекса Гольджи). В печеночных клетках гладкая ЭПС принимает участие в разрушении и обезвреживании ряда токсичных и лекарственных веществ (например, барбитуратов). В поперечно-полосатой мускулатуре канальцы и цистерны гладкой ЭПС депонируют ионы кальцин. Комплекс Гольджи- пластинчатый комплекс Гольджи что упаковочный центр клетки. Представляет собой совокупность дик тиосом (от нескольких десятков до сотен и тысяч па одну клетку). Дикгиосома— стопка из 3—12 уплощенных цистерн овальной формы, по краям которых расположены мелкие пузырьки (везикулы). Более крупные расширения цистерн дают вакуоли, содержащие резерв воды в клетке и отвечающие за поддержание тургора. Пластинчатый комплекс дает начало секреторным вакуолям, и которых содержатся вещества, предназначенные для вывода из клетки. 11ри этом просекрет, поступающий в вакуоль из зоны синтеза, (')ПС, митохондрии, рибосомы), подвергается здесь некоторым химическим превращениям. Комплекс Гольджи дает начало первичным лизосомам. В диктиосомах также синтезируются полисахариды, гликопротеиды и гликолиииды, которые затем идут на построение цитоплазматических мембран. Рибосома — важнейший немембранный органоид живой клетки сферической или слегка эллипсоидной формы, диаметром 100—200 ангстрем, состоящий из большой и малой субъединиц. Ф-ции: биосинтеза белка из а/к по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией. Рибосомы располагаются на мембранах эндоплазматического ретикулума, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Рибосомы представляют собой нуклеопротеид, в составе которого отношение РНК/белок составляет 1:1 у высших животных и 60-65:35-40 у бактерий. Рибосомная РНК составляет около 70 % всей РНК клетки. Рибосомы эукариот включают четыре молекулы рРНК, из них 18S, 5.8S и 28S рРНК синтезируются в ядрышке РНК полимеразой I в виде единого предшественника (45S), который затем подвергается модификациям и нарезанию. 5S рРНК синтезируется РНК полимеразой III в другой части генома и не нуждаются в дополнительных модификациях. Почти вся рРНК находится в виде магниевой соли, что необходимо для поддержания структуры; при удалении ионов магния рибосома подвергается диссоциации на субъединицы. Константа седиментации (скорость оседания в ультрацентрифуге) рибосом эукариотических клеток равняется 80S (большая и малая субъединицы 60S и 40S, соответственно), бактериальных клеток (а также митохондрий и пластид) — 70S (большая и малая субъединицы 50S и 30S, соответственно). Вопрос №27. Современные представления о механизме внутриклеточного транспорта. Потоки информации, энергии и веществ осуществляются непрерывно и составляют необходимое условие сохранения клетки как живой системы. Кроме структур и процессов, прямо включенных в названные потоки, в клетке функционируют механизмы, которые также являются жизненно необходимыми. Так, лизосомы, воздействуя ферментами на пиноцитированный или аутофагированный материал, обеспечивают гидролитическое расщепление макромолекул до низкомолекулярных соединений. Они же обусловливают разрушение внутриклеточных структур, утративших свое функциональное значение. Образовавшиеся при этом химические соединения включаются в потоки энергии, веществ и информации. Пероксисомы ликвидируют возникающие в клетке пероксиды, токсичные для живой протоплазмы. Организация внутриклеточных транспортных потоков обусловливается наличием и активностью микротрубочек, микрофибрилл. Вопрос №28. Внутриклеточный поток энергии Поток энергии у представителей разных групп организмов обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов концентрации веществ), электрическую, механическую, ре-гуляторную. Макроэргическим называют соединение, в химических связях которого запасена энергия в форме, доступной для использования в биологических процессах. Универсальным соединением такого рода служит АТФ. Основное количество энергии заключено в связи, присоединяющей третий остаток фосфорной кислоты. 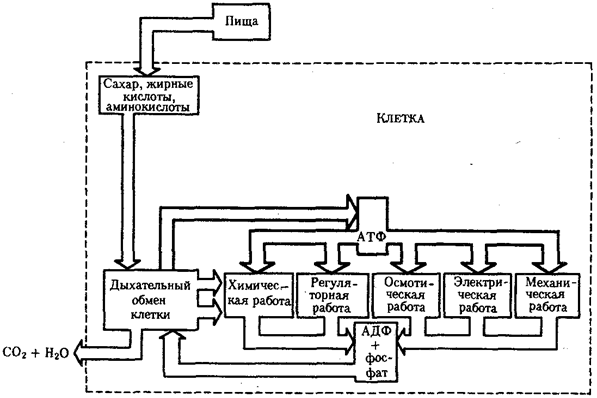 Среди органелл животной клетки особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования, а также матриксу цитоплазмы, в котором протекает процесс бескислородного расщепления глюкозы — анаэробный гликолиз (рис. 2.8). Из двух механизмов, обеспечивающих жизнедеятельность клетки энергией, анаэробный гликолиз менее эффективен. В связи с неполным (в отсутствие кислорода) окислением, прежде всего глюкозы, в процессе гликолиза для нужд клетки извлекается не более 10% энергии. Недоокисленные продукты гликолиза (пируват) поступают в митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию. Из преобразователей энергии химических связей АТФ в работу наиболее изучена механохимическая система поперечно-полосатой мышцы. Она состоит из сократительных белков (актомиозиновый комплекс) и фермента аденозинтрифосфатазы, расщепляющего АТФ с высвобождением энергии. Особенность потока энергии растительной клетки состоит в наличии фотосинтеза — механизма преобразования энергии солнечного света в энергию химических связей органических веществ. Механизмы энергообеспечения клетки отличаются эффективностью. Коэффициенты полезного действия хлоропласта и митохондрий, достигая соответственно 25 и 45—60%, существенно превосходят аналогичный показатель паровой машины (8%) или двигателя внутреннего сгорания (17%). Вопрос №29. Гипотеза эволюционного происхождения митохондрий, хлоропластов, ядра,ЭПР. Возникновение клеточной организации в ходе эволюции. На основании сходства бактерий с митохондриями и хлоропластами эукариотических клеток можно предположить, что митохондрии и хлоропласты произошли от бактерий, которые нашли себе «убежище» в более крупных гетеротрофных клетках эукариот. Бактерии имели возможность использовать молекулярный кислород для окисления питательных веществ и использовать энергию света. Более крупные клетки ? хозяева использовали эти полезные свойства и имели с такими помощниками явное преимущество перед своими современниками. Все ныне живущие эукариоты, за малым исключением, содержат митохондрии, а все автотрофные эукариоты содержат также хлоропласты. По-видимому, они были приобретены в результате независимых случаев симбиоза. Более крупные клетки эукариот защищали свои симбиотические органеллы от неблагоприятных воздействий. Этиопласты образуются у растений, выращиваемых в темноте, они имеются, например, у проростков, расположенных в почве, до их выхода на дневную поверхность. Этиопласты занимают промежуточное положение между пропластидами и настоящими хлоропластами. Для них характерно хорошо развитое проламеллярное тело с кристаллической структурой. На свету этиопласты тут же превращаются в зрелые хлоропласты. Хромопласты – это пигментированные окрашенные пластиды, но в отличие от хлоропластов, они не содержат хлорофиллов, а синтезируют и накапливают каротиноиды. Каротиноиды придают этим пластидам желтую, оранжевую и красную окраску. При этом каротиноиды синтезируются не на поверхности внутренних мембран, а в строме хромопластов. Как правило, каротиноиды растворены в жирных маслах пластоглобул. Внутренняя мембранная система у хромопластов либо не развита, либо деградировала. Форма хромопластов весьма разнообразна. Они придают яркую окраску лепесткам цветов, зрелым плодам. Это имеет явное приспособительное значение. Хромопласты обычно возникают из хлоропластов, реже из лейкопластов. По целому ряду признаков их можно назвать стареющими пластидами. Старение хлоропластов происходит, например, при созревании фруктов. Массовое старение хлоропластов наблюдается при пожелтении листьев осенью. Вопрос 30. Митохондрии. Организация потока энергии в клетке. |