Ооплазматическая сегрегация. Способы детерминации эмбриональная индукция и ооплазматическая сегрегация
 Скачать 194.54 Kb. Скачать 194.54 Kb.
|

|
Министерство образования и науки РФ ФГБОУ ВО «Бурятский государственный университет» Медицинский институт Кафедра анатомии и физиологии человека Реферат на тему: «Способы детерминации: эмбриональная индукция и ооплазматическая сегрегация" Автор: Кожинова Валентина Анатольевна, группа 141603 Улан-Удэ – 2017 ОглавлениеВведение 2 Ооплазматическая сегрегация 3 Эмбриональная индукция 6 Использованная литература: 8 ВведениеДетерминация – это путь, определяющий дальнейшее развитие частей зародыша, такое состояние клеток, когда они уже вступили на путь определенной дифференцировки, но внешне сама дифференцировка еще никак не выражается. Молекулярная природа детерминации также неизвестна, но, вероятно, включение определенных генов — если не первый, то непременный этап всякой детерминации. Существуют несколько способов детерминации:
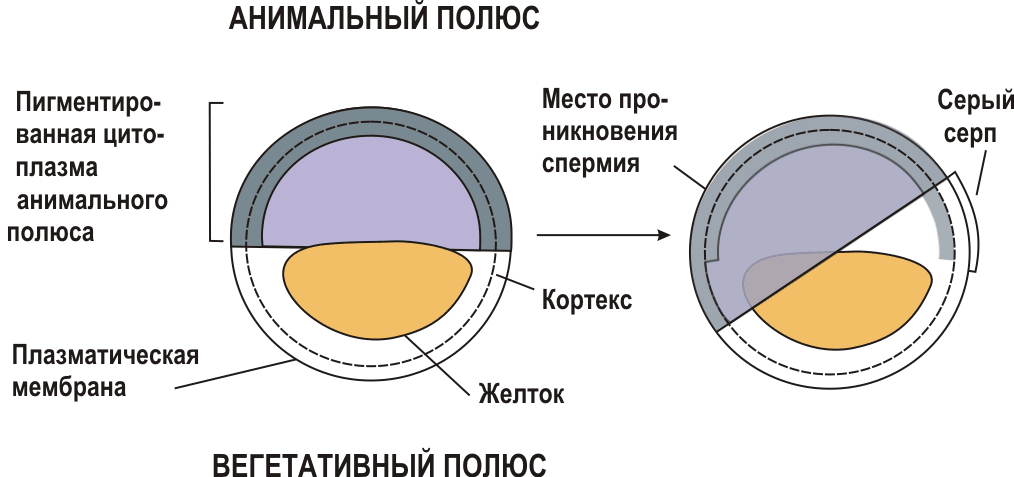
Ооплазматическая сегрегацияОоплазматическая сегрегация - возникновение различий между разными частями цитоплазмы яйца (ооплазмы), разделение (сегрегацию) яйца на зоны с несколько различными свойствами. Обычно ооплазматической сегрегацией называют те перемещения компонентов цитоплазмы, которые происходят после оплодотворения и до начала дробления яйца. В действительности же многие процессы ооплазматической сегрегации осуществляются еще в оогенезе, а некоторые продолжаются и в ходе первых делений дробления. Морской еж: Простейшая ооплазматическая сегрегация происходит в яйцах морского ежа. После четырех делений дробления бластомеры отличаются по способности образовывать некоторые структуры зародыша (султан на анимальном полюсе и кишку на вегетативном). Эта способность меняется вдоль анимально-вегетативной оси. Такое постепенное изменение свойств яйца называют градиентами. У некоторых видов морских ежей эти градиенты проявляются в виде распределения пигментных гранул, располагающихся близко к одному полюсу или образующих кольцо в области экватора. Амфибии: более сложно выглядит ооплазматическая сегрегация в яйце амфибий. Еще в ооците создается неравномерное распределение желтка и пигмента вдоль анимально-вегетативной оси: желтка больше на вегетативном полюсе, пигмент покрывает анимальную половину яйца. Ядро оплодотворенного яйца сильно сдвинуто к анимальному полюсу и окружено цитоплазмой, свободной от желтка. После оплодотворения происходит дальнейшая сегрегация: на одной из сторон яйца (она станет спинной стороной) в области экватора возникает более светлая область — серый серп (будущая хордо-мезодерма). Внутри яйца происходят и другие перемещения. В итоге в яйце амфибий можно различать три различно окрашенные зоны:
Если до оплодотворения яйцо имело только одну анимально-вегетативную ось, то теперь в нем можно провести и перпендикулярную ей дорсовентральную ось от спины к животу. Две оси образуют плоскость симметрии, которая делит будущий зародыш на правую и левую половины. Если яйцо морского ежа на стадии восьми бластомеров разделить на две или четыре части в плоскости анимально-вегетативной оси, то из каждой части, состоящей из четырех или двух клеток, разовьется нормальный зародыш. Если же такой зародыш разделить на две части по экватору, то образуются два ненормальных зародыша: один будет лишен кишки, которая образуется у вегетативного полюса, а другой — султана из длинных ресничек, который расположен с анимальной стороны. Асцидии: В яйце асцидий до оплодотворения никакой сегрегации не обнаруживается: яйцо можно разрезать пополам в любой плоскости и половина, содержащая ядро, образует нормальный зародыш. Такое яйцо изотропно. После оплодотворения в яйце происходят сложные перемещения, которые удается видеть благодаря наличию в цитоплазме различно окрашенных гранул. В итоге в яйце асцидий можно различить не менее четырех зон: 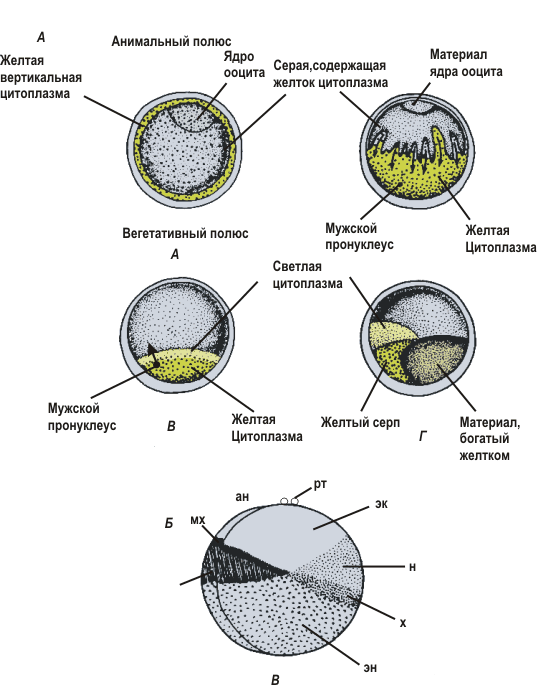
Приведенные здесь примеры относятся к тем случаям, когда за ооплазматической сегрегацией можно следить визуально благодаря различной окраске цитоплазматических компонентов. Вывод: Смысл ооплазматической сегрегации: в результате делений дробления приводит к образованию клеток, отличающихся по составу цитоплазмы. Это в свою очередь, определяет их дальнейшую судьбу. Таким образом, ооплазматическая сегрегация является первым фактором, создающим исходные различия между клетками, или, иначе, первым (по времени) фактором дифференцировки. Эмбриональная индукцияЭмбриональной индукцией называют такое влияние одной ткани на другую, соседнюю, которое вызывает в месте контакта новую дифференцировку. Иногда индуцирующая ткань (индуктор) уподобляет индуцируемую ткань себе (гомотипичная индукция). Ho в нормальном развитии индуктор вызывает в индуцируемой части второй ткани новую, третью дифференцировку. Таким образом, эмбриональная индукция приводит к увеличению числа клеточных типов и этим усиливает дифференциацию зародыша. Амфибии: в ходе гаструляции или после нее зачаток хордомезодермы вдоль спинной стороны зародыша индуцирует в эктодерме над собой зачаток нервной системы — нервную пластинку, которая свертывается в нервную трубку и дифференцируется в головной и спинной мозг. Остальная эктодерма почти це ликом становится эпителием кожи, и только на границе эктодермы и нервной пластинки узкая полоска ткани (нервный гребень) превращается в мигрирующие клетки, которые участвуют в образовании хряща и пигментных клеток кожи. Если зачаток хордомезодермы еще в самом начале гаструляции удалить, нервной системы не образуется, если же этот зачаток пересадить под раннюю эктодерму на боку или животе другого зародыша, то там образуется вторая нервная система. 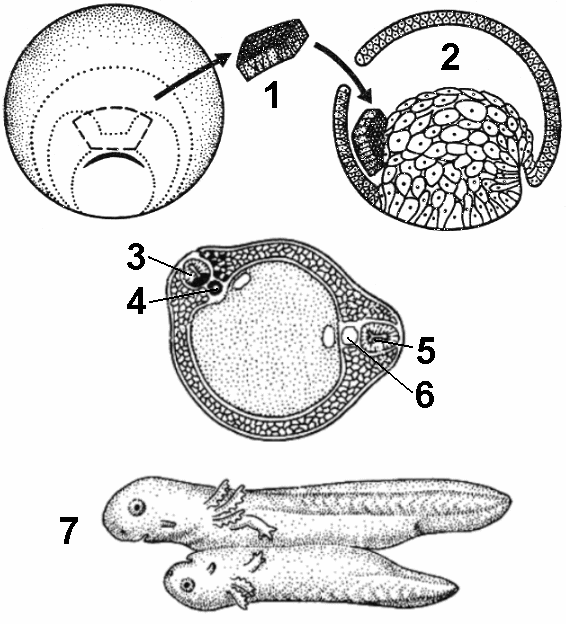 Низкая специфичность индуктора очевидна из ряда опытов. Если под эктодерму одного вида животных пересаживать индуктор от другого вида, то характер индуцируемой нервной трубки всегда идет не в соответствии с видом индуктора, а в соответствии с видовыми особенностями индуцируемой эктодермы. Индукцию можно получить при действии очень различных чужеродных источников индукции. Например, из зародыша цыпленка, причем на поздней стадии, удалось выделить индуцирующие вещества, действующие на амфибий. Все это показывает, что специфическую роль играет компетенция индуцируемых клеток — она существует только у определенной ткани и обычно только в течение короткого периода развития. Заключение: Индукция может произойти при близком контакте индуцирующей и индуцируемой тканей, во всех случаях она, по-видимому, происходит путем миграции определенных веществ. Однако в одних случаях эти вещества могут, преодолевая некоторое расстояние, мигрировать из клеток индуктора в клетки индуцируемой ткани, а в других — необходим прямой контакт между этими клетками. Вероятно, это свидетельствует о различной природе индуцирующих факторов. В недавнее время группой американских ученых был обнаружен и выделен в чистом виде индуктор, определяющий формирование головного конца животного. Индуцирующим веществом оказался олигопептид — цепочка, состоящая из одиннадцати определенных аминокислот. У морских актиний и пресноводной гидры (два разных класса) последовательность аминокислот оказалась тождественной, т. е. за много миллионов лет раздельной эволюции этих групп животных индуцирующее вещество не изменилось. Механизм индукции неясен, но трудно сомневаться в том, что в конечном итоге он состоит в активации определенных генов, запускающих всю программу дифференцировки. Роль индукции в развитии состоит в определении места дифференцировки, т. е. индуцирующее вещество — это источник позиционной информации, по которому индуцируемая клетка «узнает», где она оказалась и кем ей «следует стать». Поэтому индукцию следует считать важным (но не единственным) инструментом пространственной организации зародыша и его развития. Использованная литература:
|