Справочный материал. Глава 25 – Органы дыхания. Справочный материал по Физиологии. Глава 25 Органы дыхания
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

|
Рис. 25–8. Нервный контроль вентиляции [2]. Связи между контролирующими (нервные центры), исполнительными (дыхательные мышцы) и чувствительными (хемо- и механорецепторы) блоками даны стрелками (в том числе пунктирными для восходящей — афферентной — импульсации, а точечными для нисходящей — эфферентной — импульсации). Римскими цифрами обозначены черепные нервы (VII — лицевой, содержит двигательные, чувствительные и парасимпатические волокна; IX — языкоглоточный, повреждение нерва сопровождается нарушением глотания, анестезией верхней трети глотки, снижением нёбных и глоточных рефлексов; X — блуждающий, содержит двигательные, чувствительные и вегетативные волокна; XI — добавочный, иннервирует грудино-ключично-сосцевидную и трапециевидную мышцы; XII — подъязычный, иннервирует мышцы языка, грудино-подъязычную, грудино-щитовидную и лопаточно-подъязычную мышцы). В правой части схемы стрелки относятся к крови в кровеносной системе. Жирным прямоугольником выделены структуры ствола мозга, паренхима которого отделена от циркулирующей крови гематоэнцефалическим барьером (см. рис. 25–11). Регуляция дыхания направлена на выполнение двух задач: во-первых, автоматическая генерация частоты и силы сокращения дыхательных мышц, во-вторых, подстройка ритма и глубины дыхательных движений к реальным потребностям организма (в первую очередь, к изменениям метаболических параметров в виде Po2, Pco2 и pH артериальной крови и Pco2 и pH межклеточной жидкости мозга). Система регуляции дыхания состоит из 3 основных блоков: рецепторного (хемо- и барорецепторы, регистрирующие и передающие информацию в мозг), регуляторного, или контролирующего (совокупность дыхательных нейронов) и эффекторного (дыхательные мышцы, непосредственно осуществляющие вентиляцию лёгких). На рисунке 25–8 представлены образующие систему регуляции дыхания блоки — контролирующие (нервные центры), исполнительные (дыхательные мышцы), рецепторные (хемо- и механорецепторы) и связи между этими блоками. Таким образом, вся система регуляции дыхания состоит из нескольких взаимосвязанных регуляторных контуров. Нервные центры расположены в стволе мозга (в основном, в составе продолговатого мозга). Схема регуляции дыхания предусматривает наличие генератора ритма дыхательных движений и центра интеграции сенсорной информации. Под терминами «генератор ритма» и «интегратор сенсорной информации» следует понимать абстрагированные интегральные понятия, а не конкретные нервные структуры, так как соответствие анатомических структур рассматриваемым понятиям установлено далеко не во всех случаях. Генератор ритма включает нейроны, расположенные преимущественно в составе продолговатого мозга, а также моста и некоторых других отделов ствола мозга. Разные группы нейронов генерируют разного спектра пачки импульсов — потенциалы действия (ПД) — на разных фазах дыхательных движений, в то числе либо преимущественно во время вдоха (инспираторные нейроны), либо преимущественно во время выдоха (экспираторные нейроны). Карта расположения инспираторных и экспираторных нейронов в составе структур продолговатого мозга приведена на рисунке 25–9. 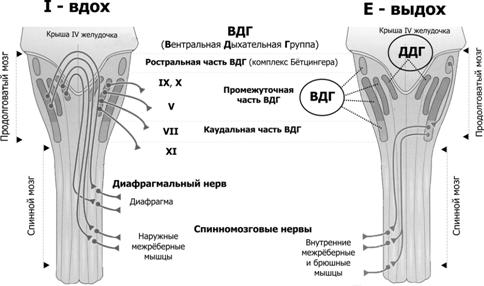 Рис. 25–9. Инспираторные (слева, Inspiration) и экспираторные (справа, Expiration) группы нейронов и их связи с дыхательными мышцами [2]. Дорсальный (сзади) вид на продолговатый и спинной мозг (мозжечок удалён). Римскими цифрами обозначены черепные нервы. Вся совокупность дыхательных нейронов подразделяется с анатомической точки зрения на вентральную и дорсальную дыхательные группы (ВДГ и ДДГ соответственно). И ВДГ, и ДДГ представлены билатерально, т.е. дублированы. Дорсальная дыхательная группа (ДДГ) содержит преимущественно инспираторные нервные клетки (в том числе нейроны важного комплекса ядер вегетативной нервной системы — ядер одиночного пути, получающих сенсорную информацию от внутренних органов грудной и брюшной полостей по нервным волокнам языкоглоточного и блуждающего нервов). Вентральная дыхательная группа (ВДГ) содержит как инспираторные, так и экспираторные нейроны. В ростро–каудальном направлении ВДГ состоит из ростральной части — комплекс Бётцингера (содержит в основном экспираторные нервные клетки и в том числе позадилицевого ядра), промежуточной (содержит преимущественно инспираторные нейроны двойного и околодвойного ядер) и каудальной (экспираторные нейроны позадидвойного ядра) частей. Направление импульсации от дыхательных нейронов: 1. от нервных клеток ДРГ к ВДГ, а также к премоторным нейронам, далее к мотонейронам и к основным инспираторным мышцам; 2. от промежуточной части ВДГ в конечном итоге к основным и вспомогательным инспираторным мышцам; 3. от каудальной части ВДГ к дополнительным экспираторным мышцам. Входящие сигналы. Генератор ритма получает нисходящую из коры головного мозга импульсацию, а также нервные сигналы от нервных клеток интегратора сенсорной информации и непосредственно от центральных хеморецепторов. Выходящие сигналы. Нервная импульсация от генератора ритма направляется к иннервирующим дыхательные мышцы двигательным нервным клеткам соответствующих ядер черепных нервов (VII, IX–XII) и к мотонейронам передних рогов спинного мозга (их аксоны в составе спинномозговых нервов направляются к дыхательным мышцам). Механизм ритмической активности генератора не установлен. Предложено несколько моделей, учитывающих индивидуальные особенности электрогенной мембраны групп однотипных нервных клеток (например, наличие разных ионных каналов), спектр синаптических связей (в том числе осуществляемых при помощи разных нейромедиаторов), наличие пейсмейкерных (со свойствами водителя ритма) дыхательных нейронов (таковые обнаружены) или пейсмейкерных свойств локальных нейронных сетей. Нет ясности и в вопросе о том, является ли ритмическая активность свойством ограниченной группы нервных клеток или свойством всей совокупности дыхательных нейронов. Интегратор сенсорной информации получает чувствительную информацию от разнообразных хемо- и механорецепторов, расположенных в органах дыхания и дыхательных мышцах, по ходу магистральных кровеносных сосудов (периферические хеморецепторы), а также в продолговатом мозге (центральные хеморецепторы). Помимо этих прямых сигналов, интегратор получает множество информации, опосредованной различными структурами мозга (в том числе и от высших отделов ЦНС). Импульсация от нервных клеток интегратора, направляясь к нейронам генератора ритма, модулирует характер разрядов от них. Чувствительные структуры, сигналы от которых прямо или опосредованно (через интегратор сенсорной информации) влияют на ритмическую активность генератора ритма, включают периферические и центральные хеморецепторы, барорецепторы стенки артерий, механорецепторы лёгких и дыхательных мышц. Наиболее существенное значение на деятельность генератора ритма имеет осуществляемый периферическими и центральными хеморецепторами контроль pH и газов крови. Периферические хеморецепторы (каротидные и аортальные тельца) регистрируют в артериальной крови pH, Po2 (Pao2) и Pco2, они особенно чувствительны к уменьшению Po2 (гипоксемии) и в меньшей степени — к увеличению Pco2 (гиперкапнии) и уменьшению pH (ацидозу). 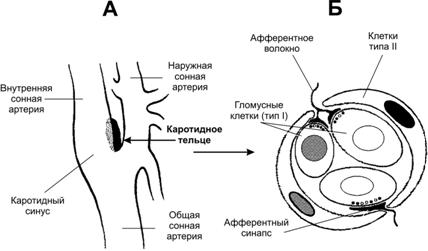 Рис. 25–10. Каротидный (сонный) синус и каротидное (сонное) тельце [11]. А. Каротидный синус — расширение просвета внутренней сонной артерии тотчас у места её ответвления от общей сонной артерии. В стенке артерии области расширения присутствуют многочисленные барорецепторы, регистрирующие значения АД и передающие эту информацию в ЦНС по нервным волокнам, проходящим в составе синусного нерва (Херинга) — ветви языкоглоточного нерва. Каротидное тельце находится в области бифуркации общей сонной артерии. Б. Клубочек каротидного тельца состоит из 2–3 клеток типа I (гломусные клетки), окружённых поддерживающими клетками (тип II). Клетки типа I образуют синапсы с терминалями афферентных нервных волокон. Каротидное тельце (рис. 25–10) состоит из скоплений клеток (клубочков, гломусов), погружённых в густую сеть кровеносных капилляров (интенсивность перфузии телец наибольшая в организме, в 40 раз больше перфузии головного мозга). Каждый клубочек содержит 2–3 хемочувствительные гломусные клетки, образующие синапсы с терминальными разветвлениями нервных волокон синусного нерва — ветви языкоглоточного нерва. В тельцах также находятся нервные клетки симпатического и парасимпатического отделов вегетативной нервной системы. На этих нейронах и на гломусных клетках заканчиваются преганглионарные симпатические и парасимпатические нервные волокна, а на гломусных клетках также постганглионарные нервные волокна из верхнего шейного симпатического узла [терминали этих волокон содержат светлые (ацетилхолин) или гранулярные (катехоламины) синаптические пузырьки]. Гломусные клетки связаны друг с другом при помощи щелевых контактов, их плазмолемма содержит потенциалозависимые ионные каналы, клетки могут генерировать ПД и содержат разные синаптические пузырьки, содержащие ацетилхолин, дофамин, норадреналин, вещество P и метионин–энкефалин. Механизм регистрации Po2, Pco2 и pH окончательно не установлен, но приводит к блокаде K+ каналов, что вызывает деполяризацию плазмолеммы гломусных клеток, открытию потенциалозависимых Ca2+ каналов, внутриклеточному увеличению [Ca2+] и секреции нейромедиаторов. Синапсы между гломусными клетками и афферентами синусного нерва. Под влиянием освобождаемых из гломусных клеток нейромедиаторов в терминалях генерируется постсинаптический потенциал и далее ПД. Эти чувствительные нейроны в свою очередь образуют синапсы с дыхательными нейронами ядра одиночного пути в составе ДДГ (см. рис. 25–9). Аортальные (парааортальные) тельца рассыпаны по внутренней поверхности дуги аорты и содержат гломусные хемочувствительные клетки, образующие синапсы с афферентами блуждающего нерва. Центральные хеморецепторы (нервные клетки стволовой части мозга) регистрируют в межклеточной жидкости мозга pH и Pco2, они особенно чувствительны к увеличению Pco2 (гиперкапнии), а часть из них к уменьшению pH (ацидозу). Существенно, что центральные хеморецепторы расположены кнутри от гематоэнцефалического барьера, т.е. они отделены от крови в системе общей циркуляции (в частности, находятся в более кислой среде). 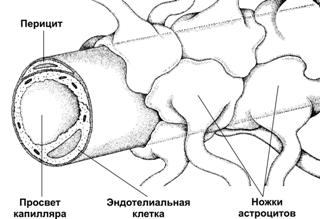 Рис. 25–11. Гематоэнцефалический барьер образован эндотелиальными клетками кровеносных капилляров мозга. Базальная мембрана, окружающая эндотелий и перициты, а также астроциты, ножки которых полностью охватывают капилляр снаружи, не являются компонентами барьера. [11]. Гематоэнцефалический барьер (рис. 25–11) изолирует мозг от временных изменений состава крови. Непрерывный эндотелий капилляров, клетки которого связаны между собой цепочками плотных контактов — основа гематоэнцефалического барьера. Гематоэнцефалический барьер функционирует как фильтр. Наибольшей проницаемостью обладают вещества нейтральные (например, O2 и CO2,) и растворимые в липидах (например, никотин, этиловый спирт, героин), но проницаемость ионов (например, Na+, Cl–, H+, HCO–3) низка. pH и Pco2. Поскольку проницаемость барьера для CO2 велика (в отличие от H+ и HCO–3), а CO2 легко диффундирует через клеточные мембраны, отсюда следует (см. уравнение 25–12), что кнутри от барьера (в интерстициальной жидкости, в ликворе, в цитоплазме клеток) наблюдается относительный ацидоз (сравнительно с кровью кнаружи от барьера, см. табл. 25–4) и что увеличение Pco2 приводит к большему уменьшению значения pH, чем в крови. Другими словами, в условиях ацидоза возрастает хемочувствительность нейронов к Pco2 и pH. Таблица 25–4. Характеристика содержания (мМ) разных компонентов в ликворе и артериальной крови [2]
Уравнение 25–12 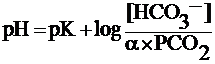 Уравнение Хендерсона–Хассельбальха рассматривает соотношение трёх переменных — рН, Pсо2, концентрации бикарбоната ([HCO3–]) — и двух констант (рК и ), где рК — обратный логарифм константы диссоциации угольной кислоты (6,1), а Чувствительные к ацидозу (хемочувствительные к Pco2 и pH) нейроны, активность которых влияет на лёгочную вентиляцию, обнаружены в вентролатеральной части продолговатого мозга, в двойном ядре, ядрах одиночного пути продолговатого мозга, а также в гипоталамусе и в голубоватом месте и в ядрах шва моста. Многие из этих хемочувствительных нейронов относятся к серотонинергическим нервным клеткам. Барорецепторы стенки артерий и вен. Эти механорецепторы реагируют на изменения давления в просвете и в стенке сосудов, они образованы терминалями волокон, проходящих в составе блуждающего и языкоглоточного нерва. Барорецепторы особенно многочисленны в дуге аорты, сонных артериях (см. рис. 25–10А), лёгочном стволе, лёгочных артериях и в стенке крупных вен большого и малого круга кровообращения. Барорецепторы участвуют в рефлекторной регуляции кровообращения и дыхания, увеличение АД может привести к рефлекторной гиповентиляции или даже остановке дыхания (апноэ), а понижение АД способно вызвать гипервентиляцию. Рецепторы воздухоносных путей и респираторного отдела регистрируют изменения объёмов лёгких, наличие посторонних частиц и раздражающих веществ и проводят информацию по нервным волокнам блуждающего и языкоглоточного (от верхних отделов воздухоносных путей) нервов к нейронам дорсальной дыхательной группы (см. рис. 25–9). К рецепторам этой группы относятся медленно адаптирующиеся рецепторы растяжения, быстро адаптирующиеся ирритантные рецепторы и J–рецепторы. Медленно адаптирующиеся рецепторы растяжения расположены среди ГМК стенки воздухоносных путей. Они реагируют на увеличение объёма лёгочной ткани (раздувание ткани лёгкого), регистрируя растяжение стенки воздухоносных путей, и проводят пачки импульсов по миелинизированным нервным волокнам. Особенность этих механорецепторов — медленная адаптируемость (при возбуждении рецепторов импульсная активность продолжается длительное время). Эти рецепторы возбуждаются при расширении просвета воздухоносных путей (бронходилатации) и запускают рефлекс Херинга–Бройера (при раздувании лёгкого происходит уменьшение дыхательного объёма и увеличение частоты дыхания; другими словами, рефлекс Херинга–Бройера направлен на подавление длительности вдоха и увеличение продолжительности выдоха). Одновременно и рефлекторно возникает тахикардия (увеличение ЧСС). У новорождённых этот рефлекс контролирует дыхательный объём при нормальном дыхании (эупноэ). У здоровых взрослых лиц рефлекс включается только при гиперпноэ — значительном увеличении дыхательного объёма (свыше 1 л), например, при значительной физической нагрузке. При обструктивных заболеваниях увеличенный объём лёгкого постоянно стимулирует рецепторы растяжения, что приводит к отсрочке следующего вдоха на фоне длительного затруднённого выдоха. Быстро адаптирующиеся (ирритантные) рецепторы расположены между эпителиальными клетками слизистой оболочки крупных воздухоносных путей. Они (как и медленно адаптирующие рецепторы растяжения) реагируют на сильное раздувание лёгочной ткани, но главным образом на действие поступающих при вдохе раздражающих ткани едких газов (например, аммиака), табачного дыма, пыли, холодного воздуха, а также на наличие в стенке воздухоносных путей гистамина (освобождается из тучных клеток при аллергических реакциях), Пг и брадикининов (поэтому их также называют ирритантными — раздражающими — рецепторами). Возбуждение от рецепторов распространяется по миелинизированным афферентным нервным волокнам блуждающего нерва. Особенность этих рецепторов — быстрая адаптируемость (при возбуждении рецепторов импульсная активность практически прекращается в течение одной секунды). При возбуждении ирритантных рецепторов увеличивается сопротивление воздухоносных путей, рефлекторно возникает задержка дыхания и кашель. J–рецепторы (от англ. «juxtacapillary» — околокапиллярные) расположены в межальвеолярных перегородках, являются как хемо– так и механорецепторами. J–рецепторы возбуждаются при перерастяжении ткани лёгкого, а также при воздействии различных экзо– и эндогенных химических соединений (капсаицин, гистамин, брадикинин, серотонин, Пг). Пачки импульсов от этих рецепторов направляются в ЦНС по безмиелиновым нервным волокнам (C волокна) блуждающего нерва. Стимуляция этих рецепторов приводит к рефлекторной задержке дыхания с последующим появлением частого и поверхностного дыхания, сужению просвета воздухоносных путей (бронхоконстрикции), увеличению секреции слизи, а также к падению АД и урежению ЧСС (брадикардии). |