лк25. Тема 5 Генеративное развитие растений. Физиология старения растений Условия перехода к репродуктивному этапу развития. Гормоны цветения
 Скачать 5.53 Mb. Скачать 5.53 Mb.
|

|
Раздел 8 Рост и развитие растений Тема 5 Генеративное развитие растений. Физиология старения растений
Переход цветковых растений от вегетативного развития и роста к генеративному развитию - сложный многофазный процесс. При достижении зрелого возраста и инициации цветения вегетативные почки преобразуются во флоральные, в которых вместо примордиев листьев закладываются зачатки органов цветка, осуществляется их формирование и рост, вслед за опылением и оплодотворением образуются семена и плоды, распространяющиеся ветром, водой, животными по окружающей территории. Полагают, что измерение времени осуществляется в организме по типу песочных часов благодаря процессам накопления растением определенной массы, числа метамеров, какого-то вещества, гормона или соотношения гормонов. Так, растения переходят к образованию цветочных зачатков только после образования определенного для каждого вида (сорта) числа листьев. Возможно также измерение времени растением по принципу маятника осциллятора, по ритмическим колебаниям каких-то физиологических процессов, которые мало зависят от внешних условий. По М.X. Чайлахяну (1988), в растениях имеются два типа регуляций, влияющих на переход растений от этапа молодости к зрелости, - автономный и индуцированный. Автономная регуляция (возрастной контроль) проявляется в переходе растений в цветочно-спелое состояние лишь в определенном возрасте. У большинства яровых зерновых цветочно-спелое состояние наступает при наличии у растений нескольких листьев. У плодовых деревьев оно наступает через 4-5 лет и более. Переход к цветению связывается с накоплением определенного количества и качества питательных веществ. По Г. Клебсу (1905), переход растений к цветению определяется отношением углерода к азоту. Зацветанию растений способствуют условия, вызывающие накопление сахаров, а вегетативное состояние поддерживают условия, способствующие накоплению азотистых веществ. У плодовых деревьев кольцевание ускоряет образование цветочных почек. В переходе к цветочно-спелому состоянию важную роль играет накопление гормональных веществ в побегах, что объясняет определенную последовательность в зацветании побегов разных ярусов. Индуцированная регуляция достижения цветочно-спелого состояния вызывается определенными внешними факторами (светом, температурой и др.). Особое значение для многих видов растений имеет воздействие температуры (яровизация) и фотопериода (фотопериодизм). Следует вспомнить о значении света для регуляции процессов жизнедеятельности растения в онтогенезе. В семени и проростке имеются системы, воспринимающие действие света разной интенсивности и качества (фотоморфогенез). При прорастании семени проросток движется к источнику света (фототропизм). Далее формируются органы автотрофного питания (фотосинтез). Закладка генеративных органов многих растений зависит от длины светового дня (фотопериодизм). В поле, теплицах и фитотронах необходимо создавать благоприятные условия для лучшей реализации потенциальной продуктивности растений, что невозможно без учета наличия сложной системы регуляции процессов их роста и развития. Цветение в ботаническом смысле представляет период в жизни растений от момента раскрывания бутона до засыхания венчика и тычинок отдельного цветка. Цветение отдельной особи длится от раскрытия первых цветков до отцветания последних. В физиологическом плане цветение рассматривается как комплекс процессов, протекающих в период от начала закладки цветочных зачатков до оплодотворения и образования зиготы. Переход покрытосеменных растений к цветению включает компетенцию, инициацию и эвокацию. Компетенция (способность зацветать) возникает у разных растений только в определенном возрасте. Так, яблоня зацветает в возрасте 5-6 лет, а дуб - свыше 40 лет. Для зацветания необходимо также, чтобы растение полностью завершило ювенильный этап онтогенеза. Большое значение имеет накопление достаточного количества питательных веществ, столь необходимых для образования репродуктивных органов и семян. Существует принцип минимального количества листьев, необходимых для заложения первых цветков. Время зацветания (переход к репродуктивной фазе) в значительной степени связано с важнейшими факторами внешней среды. Известно, что повышенное азотное питание, усиливающее вегетативный рост растений, тормозит цветение. Недостаток влаги в почве, как и недостаток многих других жизненно важных элементов, как правило, ускоряет репродуктивное развитие растений. В этом выражается стремление каждого вида создать свое потомство даже в неблагоприятных внешних условиях. Существенную роль в цветении играют некоторые микроэлементы и в первую очередь железо и медь. Наиболее продолжительное цветение наблюдается в благоприятные годы, когда температура, влажность и другие факторы находятся на уровне средних многолетних. Инициация цветения связана с восприятием растением специфических внешних и внутренних факторов, создающих условия для закладки цветочных зачатков. К важнейшим экзогенным факторам относятся чередование дня и ночи (фотопериодизм) и температурные условия, необходимые для яровизации. Эндогенные факторы связаны с возрастными изменениями растения. Эвокация цветения представляет собой завершающую фазу инициации, во время которой в апексе происходят процессы, необходимые для закладки цветочных зачатков. Процессы эти происходят на разных уровнях. На субклеточном уровне увеличивается содержание субстратов дыхания и его интенсивность, усиливается синтез РНК и белков, изменяется состав последних, повышается активность некоторых ферментов. На клеточном уровне происходит синхронизация биохимических процессов в клетках, в результате чего наступает их одновременное и ускоренное деление. На гистологическом уровне идет реорганизация меристемы: исчезают зоны, вакуолизируется стержневая сердцевинная меристема. На макроморфологическом уровне происходят ранняя инициация боковых меристем, увеличение скорости формирования бугорков, изменяется порядок расположения листьев на стебле. Таким образом, в апикальной меристеме происходят изменения, обеспечивающие развитие цветка и выполнение ими генеративных функций. В результате дифференциации тканей образуются структурные элементы цветка: чашечка, венчик, андроцей и гинецей. В пыльниках тычинок в результате последовательных редукционного и эквационного делений образуются пыльцевые зерна (мужской гаметофит). Пыльцевое зерно состоит из двух оболочек (эктина и интина), вегетативной и генеративной клеток. В завязи пестика при мейозе развивается зародышевый мешок (женский гаметофит). Он состоит из восьми ядер, дифференцирующихся затем в гаплоидную яйцеклетку, две синергиды, три антиподы и диплоидное центральное ядро. Инициация перехода к зацветанию включает в себя две фазы: индукцию и эвокацию. Индукция. Эта фаза осуществляется под действием экологических факторов - температуры (яровизация) и чередования дня и ночи (фотопериодизм) - или эндогенных факторов, обусловленных возрастом растения. Яровизация - процесс, протекающий в озимых формах однолетних и двулетних растений под действием низких положительных температур определенной длительности, способствующий последующему ускорению развития этих растений. Фотопериодизм - реакция растений на суточный ритм освещения, т.е. на соотношение длины дня и ночи (фотопериоды), выражающаяся в изменении процессов роста и развития. Одно из основных проявлений этой реакции - фотопериодическая индукция зацветания. Оба фактора -температурный и световой - могут действовать последовательно, например у озимых злаков (рожь, пшеница). Температурная и фотопериодическая регуляции служат приспособлением растений к условиям существования, так как обусловливают благоприятные сроки для перехода к зацветанию. В ходе фотопериодической индукции в листьях образуется стимулятор цветения, который транспортируется в вегетативные почки побегов, где включает вторую фазу инициации - эвокацию. Яровизация. Растения, нуждающиеся в яровизации, называют озимыми, а развивающиеся без нее - яровыми. Виды без яровизации не зацветают (качественная реакция), у других при воздействии низкими температурами лишь укорачивается время перехода к цветению (количественная реакция). У растений с количественной реакцией яровизация может проходить в наклюнувшихся семенах. Растения с качественной реакцией (двулетние и многолетние) должны достичь определенных размеров до ее начала. Яровизация обычно продолжается 1-3 месяца. Наиболее эффективны температуры от 0 до 7 °С. Для теплолюбивых растений оптимальны более высокие температуры порядка 10 - 13 °С. У некоторых видов, например у озимой ржи и капусты, периодическое прерывание низкотемпературного воздействия снижает и даже устраняет эффект яровизации. Необходимое условие яровизации - наличие делящихся клеток. Она протекает в зародыше или в апикальных меристемах стебля и в молодых листьях. Яровизация успешно осуществляется в изолированных верхушках стеблей, например у моркови и капусты. Изменения, возникшие в период яровизации, передаются только путем деления клеток. В ряде опытов показано, что непрерывный свет способствует переходу растений от озимого поведения к типичному яровому. После яровизации большинству растений необходимы длинно дневные фотопериоды. У других растений яровизация ослабляет или даже устраняет потребность в последующем фотопериодическом воздействии. Физиолого-биохимическая сущность яровизации до сих пор не ясна. У некоторых видов в ходе яровизации или после ее окончания увеличивается содержание гиббереллинов в тканях. Фотопериодизм. Понятия фотопериод и фотопериодизм были введены в науку американскими физиологами растений У. Гарнером и Г. А. Аллардом (1920-1923). В зависимости от реакции на длину дня, ускоряющей зацветание, растения делятся на длиннодневные (ДДР), короткодневные (КДР), растения, нуждающиеся в чередовании разных фотопериодов, -длинно-короткодневные (ДКДР) и коротко-длиннодневные (КДДР), а также на нейтральные по отношению к длине дня растения (НДР). К ДДР относятся, в частности, хлебные злаки, многие крестоцветные, укроп и др., к КДР - рис, соя, дурнишник, конопля, к НДР - гречиха, горох и т.д. ДДР распространены в основном в умеренных и приполярных широтах, КДР - в субтропиках. Отнесение растений к той или иной фотопериодической группе не связано с конкретной оптимальной длиной дня, а дает представление о том, ускоряется ли переход к цветению при увеличении или уменьшении длительности освещения в каждом фотопериоде. Среди растений есть виды и разновидности как с качественным, так и с количественным типами фотопериодических реакций. Интенсивность освещения, температура и другие факторы могут менять характер фотопериодических ответов растений. Для КДР с качественной реакцией решающим фактором служит длина темнового периода. Кратковременное освещение этих растений (1 мин) в середине темнового периода препятствует их переходу к цветению. Прерывание светового периода темнотой не влияет на сроки цветения. ДДР не нуждаются в периодах темноты и зацветают на непрерывном свету. Фотопериодические воздействия воспринимаются главным образом листьями, а не апексами побега. У большинства растений максимальной чувствительностью к фотопериоду обладают листья, только что прекратившие рост. Для дурнишника, зацветание которого индуцируется одним короткодневным фотопериодом, для фотоиндукции зацветания достаточно нескольких квадратных сантиметров листа. Основную роль в восприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма. 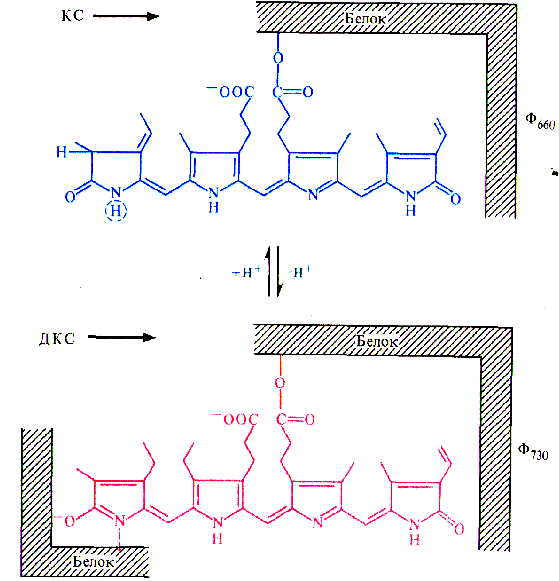 Конформационные переходы фитохрома при облучении тканей красным (КС) или дальним красным (ДКС) светом Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассимилятах. Устранение СО2 из воздуха во время светового периода снижает инициацию цветения у некоторых КДР и ДДР. У фотопериодически нейтральных растений переход к зацветанию обеспечивается возрастными изменениями (эндогенная регуляция). Эвокация (от лат. evocatio - вызывание) представляет собой завершающую фазу инициации цветения, во время которой в апексе происходят процессы, необходимые для инициации цветочных зачатков. При поступлении флорального стимула в апикальных меристемах последовательно возрастают содержание растворимых Сахаров и активность инвертазы, число митохондрий и интенсивность дыхания; активируется митоз как в периферической, так и центральной зонах стеблевой меристемы; наблюдается кратковременная синхронизация клеточных делений; усиливается синтез РНК и белков, изменяется их качественный состав (последнее - за сутки до заложения зачатков цветков). Все эти данные показывают, что сущность эвокации заключается в переключении генетической программы, способствующей развитию вегетативных почек, на другую, обеспечивающую закладку и формирование цветков. Природа флорального стимула. Немецкий ботаник Г. Клебс (1903) выдвинул теорию, согласно которой половое размножение всех групп растений (от водорослей до высших) зависит от условий питания. В его опытах такие водоросли, как Chlamydomonas, Vaucheria, и даже некоторые высшие растения (Glechomahederata), не проявляли никаких признаков перехода к половому размножению, если постоянно поддерживались благоприятные для них условия существования: состав питательной среды, освещение, температура. Изменение же этих условий в неблагоприятную для вегетативного роста сторону способствовало переходу к половому размножению. В 1913 г. Клебс высказал предположение, что цветению высших растений способствует высокое соотношение содержания эндогенных углеводов и азотистых соединений (C/N). Действительно, неблагоприятные факторы среды часто ускоряют цветение, а высокий уровень питания, особенно азотного, у некоторых растений вызывает усиление вегетативного роста и задержку генеративного развития. Однако последующее открытие фотопериодизма, выяснение роли фитохрома и фито-гормонов в процессах роста и развития показали, что регуляция перехода растений к цветению - гораздо более сложное явление. Было установлено, что фотопериодическое воздействие воспринимается листьями и затем флоральный стимул передается в апекс побега. Эти факты позволили М.X. Чайлахяну (1937) сформулировать гормональную теорию развития растений. Согласно этой теории при благоприятном фотопериоде в листьях образуется гормон цветения флориген, индуцирующий переход растений к зацветанию. Эта точка зрения подтверждалась опытами с прививками: зацветание вегетирующих растений, привитых на цветущие, происходило при неблагоприятном фотопериоде за счет веществ, передвигающихся из листьев подвоев. Химически флориген не является видоспецифичным, так как цветущие растения одного вида способны индуцировать цветение у привитых вегетирующих растений другого вида. А. Ланг (США) в 1956 г. установил, что цветение многих ДДР значительно ускоряется благодаря обработке гиббереллином. Однако в дальнейшем оказалось, что под действием ГА3 активируется прежде всего развитие цветоносов. Чайлахян (1958) выдвинул гипотезу о бикомпонентной природе флоригена, согласно которой гормон цветения состоит из гиббереллинов и антезинов, причем гиббереллины обусловливают образование и рост цветочных стеблей, а антезины индуцируют заложение цветков. По этой гипотезе ДДР, особенно розеточные формы, содержат антезины, но бедны гиббереллинами. В условиях благоприятного длиннодневного фотопериода в листьях ДДР синтезируется значительное количество гиббереллинов, которые поступают в апекс побега. Присутствие обоих компонентов флоригена - гиббереллина и антезина - индуцирует зацветание растений. 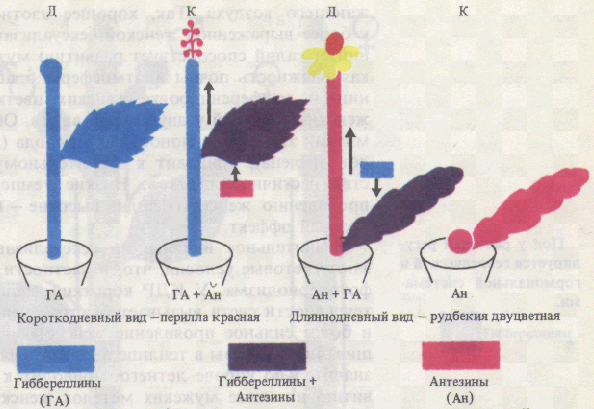 Бикомпонентная гормональная система и механизмы регуляции цветения (М. X. Чайлахян, 1978): Д - растения на длинном дне, К - на коротком дне У КДР, которые, как правило, имеют высокий стебель, наоборот, гиббереллинов достаточно при любом фотопериоде, а антезин синтезируется только в условиях короткодневного фотопериода. Этим объясняется благоприятное действие короткодневного фотопериода на зацветание КДР. У НДР содержание гиббереллинов и антезинов увеличивается с возрастом до какого-то критического уровня, который необходим для инициации цветения. По бикомпонентной гипотезе флоригена предполагается участие и других известных в настоящее время фитогормонов в регуляции закладки и формирования цветков у всех фотопериодических групп. Теория флоригена в современном ее виде достаточно убедительно объясняет механизм регуляции перехода в генеративное состояние. Однако природа антезина остается неизвестной. Ж. Бернье, Ж.-М. Кине и Р. Сакс (1985) выдвинули свою гипотезу индукции и эвокации цветения. По мнению этих авторов, эвокация контролируется не одним каким-то специфическим морфогеном, а сложной системой из нескольких факторов, каждый из которых запускает свою цепь эвокационных процессов. Взаимодействие этих процессов и приводит к закладке цветков. Факторы, участвующие в регуляции эвокации, образуются в разных частях растительного организма, причем они не обязательно идентичны у всех видов. К сказанному нужно добавить, что в листьях растений в условиях неблагоприятного фотопериода обнаруживаются ингибиторы цветения. Удаление всех листьев с одновременной подкормкой сахарозой способствует цветению некоторых ДДР и КДР в строго неиндуктивных условиях. При благоприятном фотопериоде концентрация этих ингибиторов снижается, что и приводит к зацветанию растений. Под детерминацией (определением) пола у растений понимают формирование клеток, органов или особей определенного пола в зависимости от генетических факторов, локализованных в хромосомах (генетическое определение пола), и от условий внешней и внутренней среды (фенотипическое определение пола). Цветки как органы полового размножения могут быть обоеполыми или реже раздельнополыми. Они формируются на одних и тех же (однодомность) или на разных (двудомность) растениях. Двудомных видов мало. К ним относятся конопля, хмель, тополь, ива. У некоторых двудомных видов в клетках обнаружены половые хромосомы типа XX и XY (некоторые мхи, элодея, дрема). Генетической основой определения пола у этих двудомных растений являются Х- и Y-хромосомы, однако гены, ответственные за детерминацию пола, локализуются не только в половых хромосомах, но и в аутосомах (т.е. во всех других хромосомах). Тем более это относится к растениям, у которых специализированных половых хромосом нет. Гены, ответственные за определение пола, играют важную роль, позволяя клеткам по-разному реагировать на действие факторов внутренней и внешней среды. В отличие от животных процесс формирования пола в ходе онтогенеза растений может сильно меняться в зависимости от условий вплоть до превращения одного пола в другой. Проявление пола у растений зависит от таких факторов внешней среды, как длина дня, интенсивность и спектральный состав света, температура, минеральное питание, состав окружающего воздуха. Так, хорошее азотное питание приводит к более выраженной женской сексуализации двудомных растений, а калий способствует развитию мужских растений. Высокая влажность почвы и атмосферы благоприятствует заложению и дифференцировке женских цветков и формированию женских растений у двудомных видов. Обработка такими газами, как этилен или монооксид углерода (действующий как аналог этилена), приводит к значительному увеличению количества пестичных цветков. Низкие температуры способствуют проявлению женского пола, высокие -- вызывают противоположный эффект. Значительное влияние на сексуализацию растений оказывают световые условия, что, в частности, связано с явлениями фотопериодизма. У КДР короткий день или уменьшение интенсивности света вызывает быстрое репродуктивное развитие и более сильное проявление женского пола. Например, выращивание кукурузы в теплице в осенне-зимнее время, когда день значительно короче летнего, приводит к стопроцентному развитию на месте мужских метелок женских соцветий. У некоторых растений в этих условиях усиливается женская сексуализация, у других - мужская. Характер дифференциации пола зависит не только от фотопериода, но и от спектрального состава света. Коротковолновый свет усиливает проявление женских признаков, а длинноволновый тормозит этот процесс. Регуляция пола у раздельнополых растений контролируется фитохромной системой и связана с содержанием и активностью фитогормонов. Действие факторов внешней среды в значительной степени осуществляется через изменение гормонального обмена. Исследования влияния фитогормонов и ретардантов на сексуализацию дают основание для достаточно определенных выводов о роли гиббереллинов как гормонов, усиливающих маскулинизацию (мужскую сексуализацию), и о роли цитокининов, а также ауксина и этилена как фитогормонов, усиливающих феминизацию (женскую сексуализацию) растений. В связи с изучением роли фитогормонов в проявлении пола следует подробнее остановиться на значении в этом процессе синтезирующих их органов. М. X. Чайлахян с сотрудниками (1977- 1982) показали, что как у двудомных, так и однодомных растений удаление корней, синтезирующих цитокинины, способствует усилению мужской сексуализации. 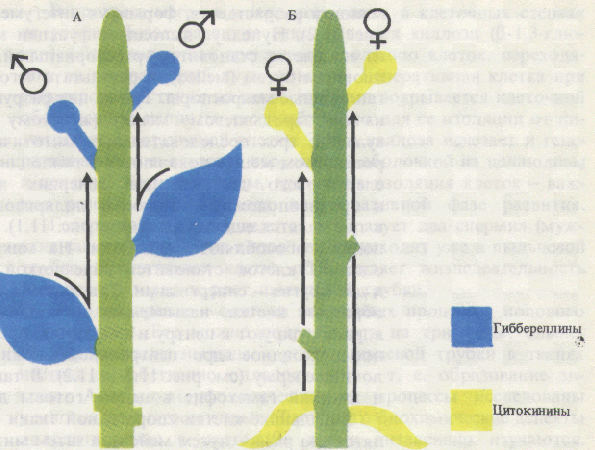 Роль органов и синтезируемых в них фито-гормонов в проявлении пола у двудомных растений (М. X. Чай-лахян, 1978). Удалены корни (А) или листья (Б) Удаление листьев приводит к противоположному эффекту, а одновременное введение гиббереллинов вновь увеличивает число мужских растений. Таким образом, корни влияют на сексуализацию растений как органы, синтезирующие цитокинины, а листья - как органы, синтезирующие гиббереллины. В связи с этим становится понятным, почему мужские экземпляры тополей при ежегодной подрезке крон (уменьшении массы листьев) превращаются в деревья с женской сексуализацией, образующие к досаде горожан большое количество тополиного пуха. Механизм действия фитогормонов на сексуализацию связан с изменением активности генного аппарата, о чем свидетельствуют опыты по изучению влияния ингибиторов синтеза нуклеиновых кислот и белка на проявление пола растений. Таким образом, процесс дифференциации пола можно представить следующим образом. Все факторы внешней среды так или иначе влияют на гормональный обмен растений, причем факторы, приводящие к сдвигу гормонального баланса в сторону увеличения содержания цитокинина (или в ряде случаев ауксина) усиливают женскую сексуализацию, а вызывающие увеличение содержания гиббереллина - мужскую. Гормональная система взаимодействует с генетическим аппаратом, что и обусловливает проявление пола у растений. |