лк25. Тема 5 Генеративное развитие растений. Физиология старения растений Условия перехода к репродуктивному этапу развития. Гормоны цветения
 Скачать 5.53 Mb. Скачать 5.53 Mb.
|

|
Формирование семян и плодов. Влияние внешних условий на цветение и плодоношение древесных растений В ходе эвокации под действием флорального стимула в апикальной меристеме стебля индуцируется экспрессия генов, ответственных за морфогенетическую программу цветообразования. Увеличивается число клеточных делений, изменяется форма апекса. Первыми закладываются чашелистики, появляются примордии лепестков, затем последовательно закладываются тычинки и плодолистики. Прежде всего, как и при формировании примордиев листа, физиологически и морфологически выделяются группы инициальных клеток, особенности деления которых определяют дальнейший ход морфогенеза каждой части цветка. Например, тычиночная нить образуется за счет апикального, а затем интеркалярного деления и роста клеток примордия. В дистальной части инициальные клетки дают начало тканям пыльника. Примордии плодолистиков приобретают подковообразную форму, удлиняются и срастаются краями. Таким образом образуется пестик. На внутренней стороне каждого плодолистика закладывается примордии семяпочки. Семяпочка разрастается, формируя интегументы и нуцеллус. Нуцеллус выполняет функции мегаспорангия. Одна из его клеток становится археспориальной. Она делится редукционным путем (мейоз), в результате чего образуются четыре гаплоидные макроспоры. Три из них разрушаются, а одна дает начало зародышевому мешку (женскому гаметофиту). В результате трех последовательных митотических делений в зародышевом мешке появляются 8 гаплоидных ядер, причем после первого деления два дочерних ядра перемещаются к противоположным полюсам поляризованного гаметофита и там делятся еще два раза. Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится яйцеклеткой (женской гаметой), а две другие - синергидами. Три ядра на халазальном полюсе образуют клетки, называемые антиподами. Два оставшихся ядра мигрируют к центру и сливаются, превращаясь во вторичное диплоидное ядро центральной клетки, которая дает начало эндосперму. В таком виде яйцеклетка и женский гаметофит в целом готовы для оплодотворения. 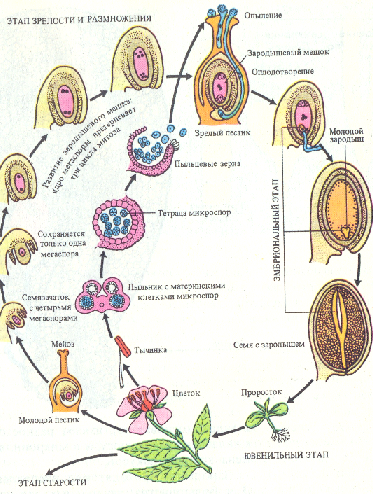 В пыльнике клетки спорогенной ткани последовательно делятся два раза — путем мейоза, а затем митоза; в результате из каждой материнской клетки образуются четыре гаплоидные микроспоры. В период образования микроспор спорогенные материнские клетки теряют большую часть ци-топлазматических РНК и белков. При прорастании ядро микроспоры делится митотически и асимметрично, в результате чего внутри пыльцевого зерна (мужского гаметофита) с вегетативным ядром появляется маленькая генеративная клетка со своим генеративным ядром. Основная роль синергид заключается в привлечении пыльцевой трубки к зародышевому мешку путем выделения хемотропных веществ и в проведении спермиев в пространство между яйцеклеткой и центральной клеткой. Возможно также, что синергиды выполняют функции гаусторий, поглощая питательные вещества из соматических клеток и снабжая ими яйцеклетку и зародышевый мешок. Роль антипод состоит в обеспечении зародышевого мешка питательными веществами. 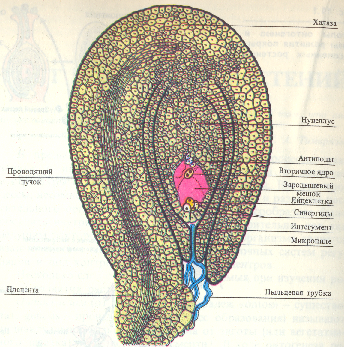 Центральный момент репродуктивного развития - индукция мейоза. Успешное завершение мейоза в культивируемых изолированных пыльниках в условиях эксперимента возможно, если споровые материнские клетки во время изоляции пыльника уже вступили в мейоз. В этом случае благотворный эффект на образование тетрад и последующие стадии развития одноядерных микроспор оказывают среды с сахарами, минеральными солями и цитокинином, гиббереллином, РНК или ее нуклеотидами. Если пыльники отрезать до начала или во время митоза пыльцевых зерен и культивировать в простой минеральной среде с добавлением сахара, то большая часть микроспор становится нежизнеспособной, у другой части вместо нормального гаметогенеза начинаются клеточные деления с образованием гаплоидных проростков (пыльцевых эмбриоидов). Опыление и оплодотворение. В процессе мега- и микроспорогенеза в клеточных стенках на ранних фазах развития откладывается каллоза, обеспечивающая временную изоляцию клеток, переходящих на генеративный путь развития. Генеративная клетка при формировании также на какое-то время покрывается клеточной оболочкой из каллозы, что необходимо для ее изоляции от цитоплазмы вегетативной клетки. Затем каллоза исчезает и генеративная клетка остается окруженной оболочкой из целлюлозы и пектина. Таким образом, временная изоляция клеток - важное условие для перехода к генеративной фазе развития. Генеративная клетка делится и образует два спермия (мужские гаметы). У многих видов это происходит уже в пыльцевой трубке. Вегетативная клетка обеспечивает жизнедеятельность пыльцевого зерна и рост пыльцевой трубки. Оплодотворение - один из важнейших процессов полового размножения. Его можно подразделить на три фазы: опыление, прорастание пыльцы и рост пыльцевой трубки в тканях пестика и собственно оплодотворение, т.е. образование зиготы. Анатомо-морфологически эти процессы исследованы у многих покрытосеменных. Физиолого-биохимические аспекты оплодотворения в настоящее время интенсивно изучаются. Для того чтобы произошло опыление, пыльца с помощью насекомых, ветра или воды должна попасть на рыльце пестика цветка. Различают пыльцевые зерна с клейкой и неклейкой поверхностью. Оболочка пыльцевого зерна состоит из внутреннего и наружного слоев: интины и экзины соответственно. Интина, помимо целлюлозы и пектиновых веществ, содержит белки, участвующие в процессах взаимного «узнавания» пыльцы и тканей рыльца, и гидролитические ферменты (кислую фосфатазу, протеазу, РНКазу и др.). Они концентрируются вблизи поры или пор прорастания и активируются при контакте поры с тканью рыльца. Экзина состоит из спорополленина - вещества терпеноидной природы и обладает высокой устойчивостью к внешним воздействиям. В полостях экзины локализованы белки, которые функционируют в системе несовместимости пыльцы и рыльца, контролируя внутривидовую и межвидовую совместимость при опылении. Пыльцевое зерно содержит ряд запасных веществ (крахмал, липиды, белки), витамины, а также активаторы и ингибиторы роста. Зрелая пыльца содержит лишь 10-15% воды. При попадании на рыльце пестика пыльца начинает набухать. Рыльца зрелых пестиков могут быть покрыты секретом или не содержат его («сухие рыльца»). Железистая ткань рыльца, выделяющая секрет, состоит из эпидермальных клеток, вытянутых в форме папилл (сосочков). Секрет вырабатывается и в субэпидермальных слоях клеток рыльца. Он состоит из липидов (производных восков), выполняющих функцию защиты от потери воды, и фенольных соединений (антоцианов, флавоноидов, коричных кислот). Фенольные компоненты могут участвовать в регуляции прорастания пыльцы, в защите от инфекций и в системе распознавания совместимости. У ряда растений секрет содержит также разнообразные белки. На рыльце одновременно может попадать много пыльцевых зерен, но прорастают лишь совместимые. Механизм несовместимости до конца не выяснен, но известно, что при контакте пыльцевого зерна с рыльцем и его набухании из экзины и интины начинают выделяться содержащиеся в них белки (гликопротеины). При контакте белков экзины с несовместимым рыльцем сразу начинается образование каллозы, изолирующей пыльцевое зерно. Прорастание несовместимой пыльцы может остановиться не только на поверхности, но и в ткани рыльца или в столбике. Так, рыльце секретирует S-факторы (гликопротеины), которые, взаимодействуя с компонентами клеточной стенки несовместимой пыльцевой трубки, подавляют ее рост. В случае совместимого рыльца активируется кутиназа и другие гидролитические ферменты интины, разрыхляющие стенки клеток рыльца и способствующие росту пыльцевой трубки. При этом начальные этапы проникновения пыльцевой трубки определяются ее положительной гидротропической реакцией, а на конечных стадиях рост пыльцевой трубки представляет собой положительную хемотропическую реакцию на вещества, выделяемые тканями микропиле, синергидами и яйцеклеткой. Сведения о веществах, необходимых для роста пыльцевых трубок, были получены при проращивании пыльцы в условиях in vitro. Пыльца большинства видов хорошо прорастает в присутствии сахарозы, различных минеральных веществ, в том числе ионов кальция; в ряде случаев требуется добавление аминокислот и гиббереллина. По-видимому, все эти соединения пыльцевая трубка получает из рыльца и из тканей столбика. Растущая пыльцевая трубка перемещается в столбике между клетками или - у некоторых видов - по пектиновым слоям в толстых клеточных стенках. Проводниковая ткань столбика способствует росту пыльцевой трубки, снабжая ее питательными веществами и активаторами роста. Пыльцевые трубки секретируют ферменты, разрыхляющие кутин рыльца и пектиновые вещества срединных пластинок клеточных стенок, что позволяет пыльцевой трубке перемещаться в столбике. Кроме того, пыльцевые трубки выделяют ауксин (или фактор, ускоряющий его синтез), участвующий в инициации развития плода. Однако резкое возрастание уровня ауксина в тканях пестика наблюдается сразу после опыления, что не может быть связано с привнесением биологически активных веществ только с пыльцой. Непосредственно после опыления наблюдается приток фосфора к завязи, изменение распределения в ней белков и углеводов. Все эти явления нельзя объяснить непосредственным воздействием пыльцевых трубок. Возможно, в регуляции подобных процессов большую роль играют электрофизиологические процессы. При опылении цветков в рыльцах появляется потенциал действия, распространяющийся в направлении завязей. ПД, по-видимому, выполняет сигнальную функцию, так как завязь отвечает на электрофизиологический сигнал изменением метаболизма, с которого начинается подготовка женской репродуктивной системы к оплодотворению. Пыльцевая трубка, пройдя через микропиле, через одну из синергид проникает в зародышевый мешок; спермин выходят и один из них сливается с ядром яйцеклетки, образуя зиготу, а другой - с полярными ядрами (или вторичным ядром) центральной клетки, дающей начало эндосперму (двойное оплодотворение, открытое в 1898 г. С.Г. Навашиным). Значение двойного оплодотворения состоит в том, что как зигота, так и первичная клетка эндосперма, имея двойную наследственность, приобретают большую жизнеспособность и приспособляемость к условиям существования. Отсюда понятна роль многочисленных приспособлений в морфологии и физиологии цветка, направленных на обеспечение перекрестного опыления. В зародышевый мешок могут проникать несколько пыльцевых трубок. Однако спермии этих трубок в оплодотворении, как правило, не участвуют и дегенерируют. Слиянию спермиев с ядрами женского гаметофита предшествует деконденсация хроматина в ядрах. Плод развивается из цветка и, как правило, содержит семена. Ткани плода формируются из завязи, но нередко в образовании плода участвуют и другие части цветка, например цветоложе, околоцветник, цветковые чешуи. Ткани плода закладываются при формировании цветка, причем в ряде случаев плод может развиваться и независимо от семян (партенокарпические, т. е. бессемянные, плоды). Однако обычно цветки без опыления и оплодотворения опадают. Развитие плодов можно разделить на четыре фазы: 1) формирование завязи до опыления; 2) рост за счет деления клеток сразу после опыления и оплодотворения; 3) рост за счет растяжения клеток; 4) созревание. Резкое усиление деления клеток завязи наблюдается вскоре после опыления. Затем наступает фаза растяжения клеток. Характер роста находится в сильной зависимости от типа плода. Сигмоидная кривая роста плодов характерна, например, для томатов, яблок, авокадо и др. В этих случаях клетки делятся почти исключительно во время формирования цветочной почки. После опыления деление клеток еще некоторое время продолжается (этот период занимает от нескольких дней у томатов до 1,5 месяцев у некоторых сортов груш). Дальше увеличение размеров плода идет за счет растяжения клеток. У плодов типа костянок (абрикос, слива, вишня и др.) рост характеризуется двойной сигмоидной кривой. Первый быстрый период роста обусловлен разрастанием завязи, нуцеллуса и интегументов семени, а зародыш и эндосперм в это время практически не развиваются. Когда же во второй фазе начинается развитие эндосперма и зародыша, завязь растет очень слабо. В это же время происходит склерификация косточки. Когда зародыш достигает полной зрелости, начинается второе ускорение роста плода, которое продолжается до полного его созревания. Таким образом, наблюдается взаимозависимость между развитием семян и ростом плодов. Деление клеток будущего плода зависит от веществ, вырабатываемых листьями в период, предшествующий цветению. Размеры меристем, обусловливающие размер плодов, сильно зависят от снабжения их углеводами и азотом, т. е. от метаболизма целого растения. Регуляция следующего этапа развития плодов связана с опылением. Одно только помещение даже чужеродной пыльцы на рыльце, без последующего оплодотворения, индуцирует разрастание стенки завязи без развития семян. Это действие можно заменить и мертвой пыльцой, и экстрактами из нее. Факторы, индуцирующие разрастание завязи и содержащиеся в экстрактах пыльцы, не являются видоспецифичными и представляют собой вещества гормональной природы. Это ауксины и гиббереллиноподобные соединения. Кроме того, при опылении повышается способность тканей пестика синтезировать собственный ауксин. Оплодотворенные яйцеклетка и эндосперм и уже начавшие развиваться семена оказывают сильное контролирующее влияние на рост плодов. Так, недоразвитие семян в силу каких-либо причин (например, воздействие паразитических организмов) служит причиной преждевременного опадения плодов. Неравномерное развитие семян в многосемянных плодах вызывает изменение их формы, развитие асимметричных, неправильной формы плодов. То, что семяпочки и развивающиеся семена регулируют рост плодов посредством вырабатываемых ими гормонов, подтверждается многими.данными. Для различных растений показано, что развивающиеся семена являются центрами синтеза ауксина, гиббереллина и цитокининов. Именно из незрелых семян впервые были выделены ГА8 и зеатин. Эти регулирующие рост вещества делают развивающийся плод центром притяжения (акцептором) питательных веществ, синтезируемых в листьях, и развитие плодов обычно связано с заметным прекращением вегетативного роста, а у однолетников - со старением всего растения. Удаление плодов задерживает старение листьев. При недостатке ауксина плоды опадают. Концепция о том, что регулирующие рост вещества, существа, существенные для роста плода (ауксин, гиббереллины, цитокинин), синтезируются в оплодотворенных семяпочках, поддерживается опытами с культурой изолированных цветков. Развитие из изолированного цветка плода до его созревания осуществляется у ряда видов на простой среде из минеральных веществ и сахарозы, если цветки были взяты спустя два дня и более после опыления материнского растения. Из цветков неопыленных томатов получали партенокарпические плоды при добавлении в культуральную среду 2-нафтоксиуксусной кислоты. Если в среду вносили томатный сок (содержащий цитокинин), то вырастали крупные плоды. Кроме перечисленных выше гормонов в плодах и семенах обнаруживаются и другие биологически активные вещества, регулирующие их развитие. Так, в плодах найдены регуляторы клеточных делений - цитокинины, а также ингибиторы роста, такие, как салициловая, кумаровая, феруловая и другие кислоты. Присутствуют также ингибиторы прорастания семян, такие, как абсцизовая кислота. Уровень и соотношение гормонов и ингибиторов от времени оплодотворения до созревания плодов сильно изменяется. Так, вскоре после оплодотворения (через неделю в завязи фасоли, например) наблюдается максимум активности гиббереллинов. Немного позже максимальной активности достигают ауксины. Увеличение содержания ауксина совпадает с переходом эндосперма к клеточному делению. С момента достижения максимума концентрации ауксина начинает активно расти зародыш. Следующий пик активности ауксина связан с усилением клеточных делений на периферии эндосперма, где и содержится максимальное количество ауксина в уже сформировавшихся семенах. Наивысший уровень регуляторов клеточных делений характерен для очень молодых плодов и семян. Созревание плодов сопровождается усиленным продуцированием этилена, который резко ускоряет эту завершающую фазу их развития. При развитии плодов и семян происходят значительные метаболические изменения. Так, дыхание плодов, очень интенсивное на первых этапах развития, снижается по мере их роста и кратковременно усиливается у многих сочных плодов в период созревания (климактерический период). Основными метаболитами, потребляемыми плодами в процессах дыхания, являются сахара и органические кислоты. Их содержание в ходе развития плодов сильно меняется. Уже через несколько дней после опыления или обработки ауксином в плодах резко снижается концентрация сахаров. Во время фаз быстрого развития плодов и семян идет интенсивный синтез крахмала, гидроли-зуемого в период созревания. В плодах некоторых растений крахмал вовсе отсутствует. В разных плодах обнаруживаются также глюкуроновая кислота, сорбитол и др. Углеводы частично синтезируются в хлоропластах, которые присутствуют в незрелых плодах, однако в основном ассимиляты поступают в плоды из фотосинтезирующих листьев. Среди органических кислот в плодах преобладают ди -и трикарбоновые, такие, как лимонная, яблочная, винная и др. В большом количестве в молодых плодах обнаруживаются ароматические кислоты. Во время роста плодов содержание органических кислот увеличивается, а при их созревании опять уменьшается. Показано, что накопление органических кислот зависит от климатических факторов: при солнечной погоде органические кислоты используются на дыхание, при пасмурной - накапливаются. Вопрос об образовании органических кислот в плодах до конца не ясен. По некоторым данным, уменьшение листовой поверхности ведет к снижению содержания кислот в плодах, что может свидетельствовать о синтезе их в листьях с последующим перемещением в плоды. По другим данным, незрелые плоды способны к темновой фиксации СО2, который включается в яблочную, лимонную и другие кислоты. Процессы созревания начинаются, когда плоды заканчивают свой рост. Созревание сочных плодов характеризуется снижением соотношения кислот и сахаров, образованием ароматических веществ, разложением хлорофилла и дубильных веществ, накоплением антоцианов и других вакуолярных пигментов, уменьшением твердости и упругости тканей благодаря гидролизу пектиновых веществ клеточных стенок. Однако на начальных этапах созревания в плодах еще преобладают процессы синтеза веществ. После прохождения периода созревания плод вступает в фазу отмирания. Превращения веществ при созревании семян. При переходе растений к этапу половой зрелости и размножения, в течение которых формируются генеративные органы, существенно изменяется ход биохимических процессов в растении. Основным аттрагирующим центром в этот период онтогенеза становятся семена, в которых протекают реакции новообразования структурных элементов и запасных веществ. В целом растении для формирования семян происходит мобилизация веществ, накопленных ранее в разных его органах. Важную роль в воспроизводстве потомства растений играют запасные вещества семени, которые обеспечивают питание проростка на самом начальном этапе онтогенеза в гетеротрофный период его развития. Основными запасными веществами семян большинства видов растений являются белки, углеводы и липиды. Распределение этих веществ в разных частях плода неодинаковое. Например, весь крахмал пшеничного зерна, 80 % сахара и больше половины белка содержатся в эндосперме. Алейроновый слой также богат питательными веществами. В нем находятся более половины имеющегося в зерновке жира и пятая часть белка и сахара. Масса зародыша составляет незначительную часть массы семени (у пшеницы около 3 %), однако концентрация веществ в нем довольно высокая. В семенах злаков имеются также свободные аминокислоты, нуклеиновые кислоты, ферменты, витамины и другие физиологически активные вещества. В золе зерна содержится большое число макро- и микроэлементов (Р, К, Mg, Al, Са, Si, Cl, Сu, Со и др.). Биологический синтез органических веществ в клетках обеспечивается в первую очередь поступлением и распределением двух основных органогенов - азота и углерода. В онтогенезе растительного организма отложение запасных азотистых соединений имеет видовую специфику. У бобовых существенную часть аминного азота, используемого для синтеза белка в формирующемся зародыше, составляют аминокислоты, содержащиеся в плодах, а также накопившиеся в самом эндосперме. У большинства же растений источником азота для образования запасного белка служат аминокислоты, поступающие из листьев, в которых активизируются процессы гидролиза азотистых веществ. Важнейший запасной продукт многих растений - крахмал - образуется в пластидах семени вследствие утилизации поступающих из окружающей цитоплазмы простых сахаров. Этот процесс катализируют три фермента: -глюканфосфорилаза, крахмалсинтетаза и Q-энзим. Первые два фермента контролируют синтез линейных цепей полисахарида, в результате чего образуется его амилозный компонент, который обусловливает ветвление этих цепей с образованием амилопектина. Исходя из особенностей синтеза крахмала в семенах, а также образования и функционирования пластид выделяют два типа семян. К первому типу относят семена, накапливающие крахмал в течение всего или большей части периода их формирования и содержащие крахмальные зерна в зрелом состоянии (многие растения семейств Бобовые, Мятликовые, Гречишные). Семена второго типа образуют крахмал на ранних этапах своего формирования до начала накопления запасных белков и липидов. В зрелом состоянии крахмал они не содержат. К ним относятся семена высокомасличных растений семейств Капустные, Молочайные Астровые. Липиды локализуются в различных частях семени - эндосперме, осевой части зародыша, семядолях. Они начинают накапливаться на очень ранних этапах развития плодов. В зародышах семян липиды в отличие от крахмала накапливаются в значительных количествах. Одновременное присутствие в эмбриональных тканях жира и крахмала - явление чрезвычайно редкое, оно отмечено у некоторых представителей гвоздичных, лавровых и др. Важно отметить, что особенно активный синтез липидов наблюдается в растениях с вступлением их в генеративную фазу развития. В семенах как основном аттрагирующем центре процессы липидных реакций протекают значительно интенсивнее, чем в вегетативных органах растений. Важным запасным веществом семян является фитин, представляющий собой кальциево-магниевую соль инозитфосфорной кислоты. Физиологические функции фитина в жизнедеятельности растений довольно обширны. Он является основным фосфогеном растений и служит резервом фосфора в зрелых семенах, который используется проростком в период гетеротрофного питания. При распаде фитина в прорастающих семенах катионы калия, кальция и магния переходят в легкотранспортируемые формы, что способствует их быстрой мобилизации в осевую часть проростка. Миоинозит - продукт гидролиза фитина - участвует в углеводном обмене. В зрелых семенах фитин находится исключительно в алейроновых зернах в форме калиево-магниево-кальциевой соли. Большая часть его содержится в сложных алейроновых зернах — глобоидах, однако некоторое его количество имеется непосредственно в белковом матриксе алейронового зерна. В семенах масличных растений фитина больше (до 3 %), чем в белково-крахмальных и крахмалистых семенах (0,3-1,5 %). Вегетативные органы растений играют важную роль в синтезе и накоплении запасных веществ в плодах и семенах. Установлено, что большая часть белка в семенах (около 2/3) синтезируется в результате реутилизации азотистых веществ вегетативных органов, где они накопились к началу налива семян. Различные органы играют неодинаковую роль в снабжении формирующегося семени азотистыми веществами. В опытах с пшеницей установлено, что из всего азота, поступающего в семена из вегетативных органов, на листья приходится около 50 %, на стебли – 20-30, на корни – 10-30 %, элементы соцветия дают 10-15 %. Вегетативные органы являются также основным источником углеводов для синтеза в зерновках крахмала. Если учесть потери на дыхание, то реальный вклад листьев в накопление в семенах углеводов составляет 110- 120 % их общего количества в зерне. Значительная роль в формировании семян принадлежит аттрагирующей способности колоса, т.е. привлечению метаболитов из вегетативных органов для собственного формирования. Колосовые, цветковые чешуи и ости наряду с высокой фотосинтетической деятельностью более активно используют мономерные вещества других органов для синтеза запасных веществ семян. У сортов с повышенной аттрагирующей способностью более высокая транспирация за счет хорошо развитых колосковых, цветковых чешуи и остей. Поступление веществ в зерновку продолжается вплоть до середины восковой спелости (влажность 30-27 %). Следовательно, это время необходимо отнести к оптимальному сроку начала раздельной уборки колосовых культур. Более ранняя уборка приводит к недобору урожая. Завершать скашивание в валки необходимо в конце восковой спелости при влажности зерна 23- 22 %. В накоплении веществ зародышем отмечаются такие же закономерности, как и при формировании зерновки в целом, причем увеличение массы зародышей имело место также и при дозревании растений при суховее, что указывает на активное использование ими запасных веществ эндосперма. Этому способствуют более высокая влажность эмбриональных тканей (75-40 % при разных сроках уборки) по сравнению с эндоспермом (59-35 % в те же сроки уборки). Если накопление веществ в зародыше продолжается вплоть до конца восковой спелости зерна, то количество клеток в нем уже после тестообразного состояния не изменяется. В зависимости от условий дозревания масса зародышей варьировала в значительных пределах, однако число клеток в них также практически не изменялось. Соотношение вегетативной массы и массы семян растения в значительной степени определяет их белковость. Как правило, у высокобелковых сортов пшеницы на единицу массы семян приходится больше вегетативной части. Основным фактором, определяющим уровень накопления белка в семенах, является количество азота в растении, приходящееся на единицу массы зерна, называемое показателем обеспеченности зерна азотом. Между соотношением развития корневой системы и надземной массы существует тесная взаимосвязь. При увеличении площади питания число корешков на одно растение возрастает. Однако при этом значительно усиливается кущение растений, образуется подгон с неполноценными зерновками, что приводит к снижению массы 1000 семян и ухудшению их биологических свойств. Причина заключается в том, что при увеличении общего количества корешков, приходящегося в целом на растение, их число в пересчете на отдельные стебли и семена уменьшается, что обусловливает снижение обеспеченности последних влагой и элементами питания. Поэтому лучшие семена получают при оптимальных площадях питания растений. Влияние внешних условий на цветение и плодоношение древесных растений. На процесс цветения сильное влияние оказывают и внешние условия - условия минерального питания, избыток азотистых соединений в почве удлиняет фазу вегетативного роста, а высокий уровень фосфорного питания, наоборот, ускоряет переход растения в репродуктивную фазу развития. Ускорению развития растений способствует сухость воздуха при достаточно высокой влажности почвы, а также освещенности. Такие условия создаются, к примеру, на орошаемых землях в Средней Азии, благодаря чему урожайность растений здесь очень высока. Все перечисленные выше факторы (освещенность, водный режим, условия минерального питания, температура воздуха и почвы) значительно влияют на репродуктивную деятельность древесных растений. Условия внешней среды воздействуют не только на сроки и обилие цветения и плодоношения, но и на проявление сексуализации побегов, соотношение мужских и женских цветков. Сильное влияние на развитие растений оказывает продолжительность дневного освещения. Фотопериодическая реакция имеет особое значение в жизни многолетних древесных растений. Фотопериод лежит в основе ритмики роста деревьев, опадения листьев, формирования и покоя почек, начала цветения и других явлений. Определяющим моментом здесь выступает продолжительность темных ночных часов. Прекращение роста в высоту при уменьшении длины дня у деревьев умеренного климата является приспособлением к внешней среде, древесный организм переходит к состоянию покоя, в котором он становится более устойчивым к неблагоприятным воздействиям еще задолго до начала первых осенних заморозков. С помощью регулирования фотопериода появилась возможность ускорения развития древесных растений. Так, удалось получить цветки и плоды с полностью созревшими семенами у акации белой, бархата амурского в более северных для них районах путем искусственного уменьшения длины дня. Этот прием играет важную роль в интродукции древесных растений. Следует отметить и явления, связанные с влиянием температуры на развитие растений. В течение длительной эволюции некоторые растения приспособились к определенной смене тепла и холода. Так, озимые злаки для нормального развития требуют, чтобы в течение 1 - 2 месяцев они находились при температуре от + 2 до + 10°С. Только после этого они развиваются нормально, цветут и дают урожай зерна. В противоположность озимым яровые злаки не требуют пониженных температур. Холод способствует переходу зимующих растений от состояния вегетативного роста к репродуктивному развитию. Данное явление, получившее название яровизации имеет значение не только для озимых зерновых, но и для двухлетних и многих многолетних растений. В настоящее время ставится на очередь проблема, связанная с действием на развитие растений повышенной температуры. Некоторые растения южного происхождения требуют для осуществления нормального хода развития воздействия высокой температуры (хлопчатник, просо, соя и др.). Молодые растения дуба, выросшие из желудей, подвергнутых такой обработке, начинали плодоносить на седьмой год жизни вместо обычных 15-20 лет. |