Биология_лекции. Тема Сущность и субстрат жизни. Свойства живого. Уровни организации живой материи. Типы клеточной организации
 Скачать 5.67 Mb. Скачать 5.67 Mb.
|

|
Сравнение растительной и животной клетки Общие признаки
Обмен веществ и энергии Обмен веществ и энергии — это важнейшее свойство живого, проявляющееся на разных уровнях организации живого. Благодаря обмену веществ и энергии происходят рост и размножение, формируются другие важнейшие свойства клеток и организмов. Обмен веществ и энергии (метаболизм) — совокупность химических реакций, протекающих в клетках или в целостном организме и заключающихся в синтезе сложных молекул и новой протоплазмы (анаболизм) и в распаде молекул с освобождением энергии (катаболизм). Энергия необходима для:
Энергетические процессы у всех живых существ сходны. В основе регуляции метаболических путей лежат общие механизмы. анаболизм и катаболизм Основные метаболические процессы:
Анаболизм (от греч. anabole — подъём), ассимиляция, совокупность химических процессов в живом организме, направленных на образование и обновление структурных частей клеток и тканей. Заключается в синтезе сложных молекул из более простых с накоплением энергии. Процессы:
Катаболизм, или диссимиляция (от лат. dissimilis — расподобление), является экзотермическим процессом, при котором происходит распад веществ с освобождением энергии. Этот распад происходит в результате переваривания и дыхания. Катаболизм (от греч. katabole — сбрасывание, разрушение), диссимиляция, совокупность ферментативных реакций в живом организме, направленных на расщепление сложных органич. веществ — белков, нуклеиновых кислот, жиров, углеводов, поступающих с пищей или запасённых в самом организме (жиры, крахмал гликоген и др.). Процессы:
В процессе катаболизма энергия, заключённая в химических связях крупных органических молекул, освобождается и запасается в форме связей АТФ. АТФ (аденозинтрифосфорная кислота). Молекулапредставляет собой нуклеотид. АТФ — главный универсальный поставщик энергии в клетках всех живых организмов. Цикл АТФ-АДФ является основным механизмом обмена энергии в живых системах. Строение:
Фосфатные группы в молекуле АТф соединены между собой высокоэнергетическими (макроэргическими) связями 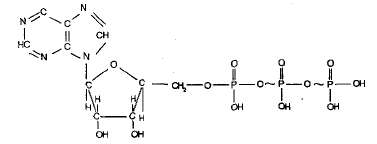 В результате гидролитического отщепления от АТФ фосфатной группы образуется АДФ (аденозиндифосфорная кислота) и высвобождается порция энергии: АТФ + Н2О АДФ +Н3РО4 + 40 кДж АДФ также может подвергаться дальнейшему гидролизу с отщеплением еще одной фосфатной группы и выделением второй порции энергии; при этом АДФ преобразуется в аденозин-монофосфат (АМФ), который далее не гидролизуется: АДФ + Н2О АМФ +Н3РО4 + 40 кДж Фосфорилирование процесс образования АТФ из АДФ и неорганического фосфата за счет энергии, освобождающейся при окислении органических веществ и в процессе фотосинтеза. При этом должно быть затрачено не менее 40 кДж/моль энергии, которая аккумулируется в макроэргических связях: АДФ+ Н3РО4+ 40 кДж АТФ + Н2О АТФ быстро обновляется. Синтез АТФ осуществляется главным образом в митохондриях и хлоропластах Образовавшаяся здесь АТФ направляется в те участки клетки, где возникает потребность в энергии. Пример: У человека каждая молекула АТФ расщепляется и вновь восстанавливается 2 400 раз в сутки, так что ее средняя продолжительность жизни менее 1 мин. АТФ не единственное биологически активное соединение, содержащее пирофосфатные связи. Некоторые фосфорилированные соединения по количеству энергии, заключенной в таких связях, не отличаются от АТФ. Однако дифосфаты таких соединений не могут заменить аденозиндифосфорную кислоту в тех процессах, которые ведут к синтезу АТФ, а их трифосфаты не могут заменить АТФ в последующих процессах энергетического обмена, в которых АТФ используется как донор энергии, необходимой для протекания биосинтетических реакций. Возможно, что такая высокая степень специфичности отражает не столько уникальность АТФ, сколько уникальные особенности биохимических процессов, приспособленных исключительно к АТФ. Поступление веществ в клетки Благодаря содержанию растворов солей, сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления. Например: давление в клетках животных (морских и океанических форм) достигает 30 атм и более. В оптимальных условиях осмотическое давление клеточного сока наземных органов болотных растений колеблется от 2 до 16 ат, у степных — от 8 до 40 ат. В разных клетках растения О. д. может резко различаться (так, у мангровых О. д. клеточного сока около 60 ат, а О. д. в сосудах ксилемы не превышает 1—2 ат). У гомойосмотических организмов, т. е. способных поддерживать относительное постоянство О. д., средней величины и диапазон колебаний О. д. различны (дождевой червь — 3,6—4,8 ат, пресноводные рыбы — 6,0—6,6, океанические костистые рыбы — 7,8—8,5, акуловые — 22,3—23,2, млекопитающие — 6,6—8,0 ат). У млекопитающих О. д. большинства биологических жидкостей равно О. д. крови (исключение составляют жидкости, выделяемые некоторыми железами, — слюна, пот, моча и др.). О. д., создаваемое в клетках животных высокомолекулярными соединениями (белки, полисахариды и др.), незначительно, но играет важную роль в обмене веществ Градиент концентрации - разность концентрации веществ внутри и снаружи клетки Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией. Основные пути поступления веществ в клетки:
Экзоцитоз —процесс секретирования клетками различных веществ Различают:
Клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом. Пассивный транспорт. Когда транспортируются незаряженные молекулы, то пассивный транспорт определяется только градиентом концентрации, т. е. разностью концентрации вещества на разных сторонах мембраны. Если же молекулы транспортируемого вещества заряжены, то к влиянию градиента концентрации добавляется влияние электических потенциалов по обе стороны мембраны. Градиент концентрации и электрический градиент в совокупности составляют электрохимический потенциал, который позволяет транспорт в клетку только положительно заряженных ионов. Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде). Катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью ферментов. Как и обычная, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ. Примеры Такие вещества, как глюкоза, почти нерастворимы в липидах, и размеры их частиц больше 0,8 нм; тем не менее они довольно быстро проходят через плазматическую мембрану, например через оболочку эритроцита. По-видимому, это обусловлено «облегченной диффузией» — присоединением к специфической молекуле-переносчику, представляющей собой пептид или белок. Молекула глюкозы G соединяется с молекулой-переносчиком X у наружной поверхности мембраны, и образовавшийся комплекс GX, растворимый в липидах, может диффундировать через мембрану к ее внутренней стороне, где он диссоциирует, и освобожденная глюкоза оказывается внутри клетки. Затем переносчик диффундирует обратно к наружной поверхности и сразу же может присоединить к себе другую молекулу глюкозы. При такой системе максимальная скорость переноса глюкозы определяется общим числом молекул-переносчиков, имеющихся в мембране, и возможными скоростями образования и расщепления комплекса GX. Для подобного процесса характерна «кинетика насыщения»: при малой концентрации глюкозы в наружном растворе скорость ее проникновения в клетку пропорциональна этой концентрации; однако при более высоких концентрациях пропорциональность исчезает, так как все молекулы переносчика уже «насыщены» глюкозой. Переносчики специфичны — они могут присоединять только глюкозу и некоторые очень близкие к ней по структуре сахара. Молекулы сахаров, сходные по своему химическому строению, будут конкурировать между собой за связывающие участки в молекулах-переносчиках. Облегченная диффузия не требует расхода энергии, если наружная концентрация глюкозы выше внутренней и глюкоза, таким образом, перемещается «вниз» по химическому градиенту. Однако некоторые клетки, например клетки кишечного эпителия и внутренней выстилки почечных канальцев, способны концентрировать глюкозу, заставляя ее перемещаться «вверх» по химическому градиенту (или, как говорят, против градиента концентрации), и для этого уже требуется затрата энергии. Гормон инсулин резко усиливает поглощение глюкозы скелетными мышцами и некоторыми клетками нашего организма. Пока еще не ясно, вызывает ли он увеличение числа эффективных молекул-переносчиков или же просто ускоряет реакции, с которыми связано образование и расщепление комплекса переносчика с глюкозой. Полагают, что не только облегченная диффузия, но и активный перенос глюкозы происходят с участием специфического переносчика, образующего с глюкозой комплекс, синтез или расщепление которого требует затраты энергии. Из клеточных мембран кишечной палочки Escherichia coli был выделен и частично очищен липопротеид, который, по-видимому, служит переносчиком, или пермеазой, для лактозы. На одну клетку приходится около 9000 молекул пермеазы. Как полагают, этот белок присоединяет лактозу у наружной поверхности мембраны, а затем образовавшийся комплекс диффундирует к ее внутренней поверхности, где лактоза отделяется от переносчика. Этот гипотетический механизм в принципе сходен с предполагаемым механизмом натриевого насоса. 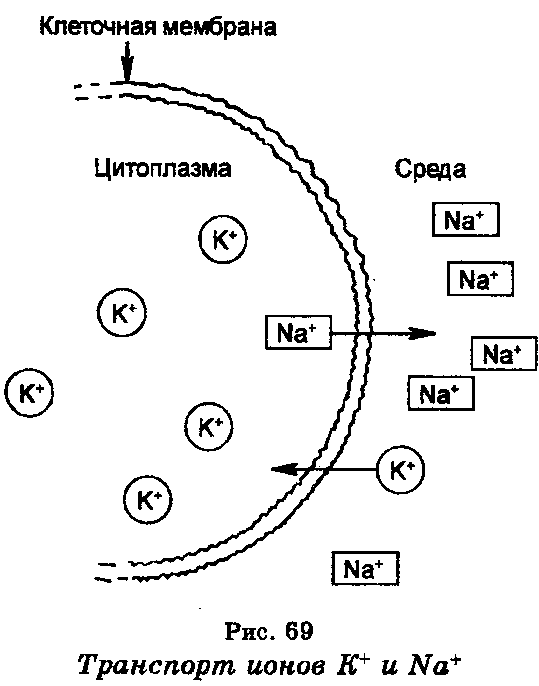 |