Активный транспорт.
Пример:
транспорт ионов натрия и калия, который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Система Na+ + К+ —АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+ внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+ сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+, благоприятный для транспорта ионов К+ и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+ из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Экзоцитоз
Конститутивный экзоцитоз осуществляется периодически, по мере накоплениях секрета, без видимого воздействия внеклеточных факторов, при колебаниях гидратированности клетки, без участия аппарата Гольджи и Са2+—независимо (Инсулин и ряд других гормонов, нейротрансмиттеров и многих ферментов.)
Нейромедиатор (Neurotransmitter) - химический посредник, освобождающийся из пресинаптического нервного окончания и передающий нервный импульс в синапсе постсинаптичсскому окончанию, мышечному волокну или железе, которые эти нервы иннервируют. Основными нейромедиаторами в периферической нервной системе являются ацетилхолин и норадреналин (сскретируются нервными окончаниями симпатической нервной системы). В центральной нервной системе наряду с ацетилхолином и норадреналином, нейромедиаторами являются дофамин, серотонин, гаммааминобутировая кислота и некоторые другие вещества.
Регулируемый экзоцитоз отличается от предыдущего необходимостью для запуска внеклеточного воздействия и четкой стадийностью процесса. Каждая из стадий регулируется дифференцированно. Специфика экзоцитоза определяется химической природой выделяеммых веществ, невезикулярной или везикулярной формами его выведения.
С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
Фотосинтез
Фотосинтез — это синтез органических соединений в листьях зеленых растений из воды и углекислого газа атмосферы с использованием солнечной (световой) энергии, адсорбируемой хлорофиллом в хлоропластах.
Благодаря фотосинтезу происходит улавливание энергии видимого света и превращение ее в химическую энергию, сохраняемую (запасаемую) в органических веществах, образуемых при фотосинтезе.
Датой открытия процесса фотосинтеза можно считать 1771 г. Английский ученый Дж. Пристли обратил внимание на изменение состава воздуха вследствие жизнедеятельности животных. В присутствии зеленых растений воздух вновь становился пригодным как для дыхания, так и для горения. В дальнейшем работами ряда ученых (Я. Ингенгауз, Ж. Сенебье, Т. Соссюр, Ж.Б. Буссенго) было установлено, что зеленые растения из воздуха поглощают С02, из которого при участии воды на свету образуется органическое вещество. Именно этот процесс в 1877 г. немецкий ученый В. Пфеффер назвал фотосинтезом. Большое значение для раскрытия сущности фотосинтеза имел закон сохранения энергии, сформулированный Р. Майером. В 1845 г. Р. Майер выдвинул предположение, что энергия, используемая растениями, — это энергия Солнца, которую растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
Основная роль фотосинтезирующих организмов:
1) трансформация энергии солнечного света в энергию химических связей органических соединений;
2) насыщение атмосферы кислородом;
В результате фотосинтеза на Земле образуется 150 млрд. т. органического вещества и выделяется около 200 млрд. т свободного кислорода в год. Он препятствует увеличению концентрации CO2 в атмосфере, предотвращая перегрев Земли (парниковый эффект).
Созданная фотосинтезом атмосфера защищает живое от губительного коротковолнового УФ-излучения (кислородно-озоновый экран атмосферы).
В урожай сельскохозяйственных растений переходит лишь 1-2% солнечной энергии, потери обусловлены неполным поглощением света. Поэтому имеется огромная перспектива повышения урожайности благодаря селекции сортов с высокой эффективностью фотосинтеза, созданию благоприятной для светопоглощения структуры посевов. В связи с этим особенно актуальными становятся разработка теоретических основ управления фотосинтезом
Значение фотосинтеза гигантское. Отметим лишь, что он поставляет топливо (энергию) и атмосферный кислород, необходимые для существования всего живого. Следовательно, роль фотосинтеза является планетарной.

Планетарность фотосинтеза определяется также тем, что благодаря круговороту кислорода и углерода (в основном) поддерживается современный состав атмосферы, что в свою очередь определяет дальнейшее поддержание жизни на Земле. Можно сказать далее, что энергия, которая запасается в продуктах фотосинтеза, есть по существу основной источник энергии, которым сейчас располагает человечество.
Суммарная реакция фотосинтеза
СО2 +Н2О = (СН2О) + О2.
Химию фотосинтеза описывают следующими уравнениями:
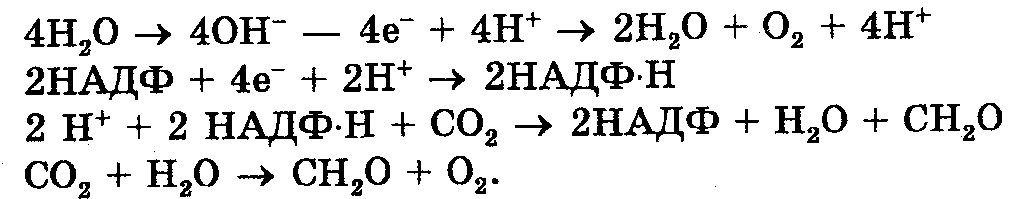
Фотосинтез – 2 группы реакций:
световая стадия(зависят отосвещенности)
темновая стадия (зависит от температуры).
Обе группы реакций протекают одновременно
Фотосинтез происходит в хлоропластах зеленых растений.
Фотосинтез начинается с улавливания и поглощения света пигментом хлорофиллом, содержащимся в хлоропластах клеток зеленых растений.
Этого оказывается достаточно, чтобы сместить спектр поглощения молекулы. 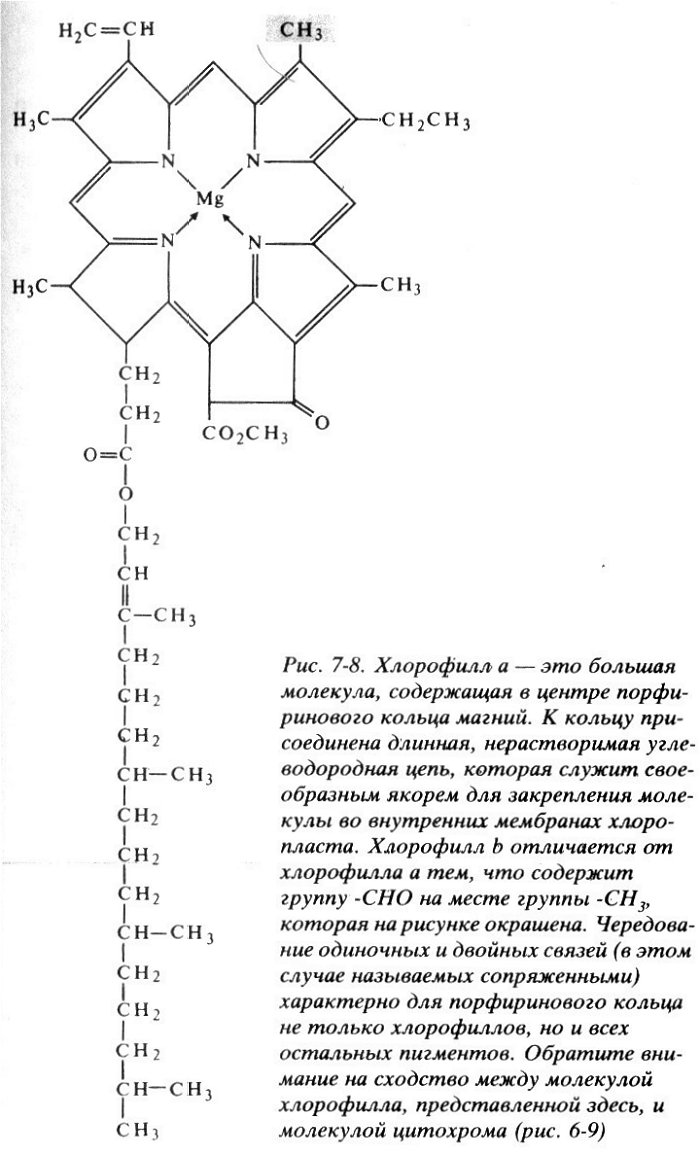
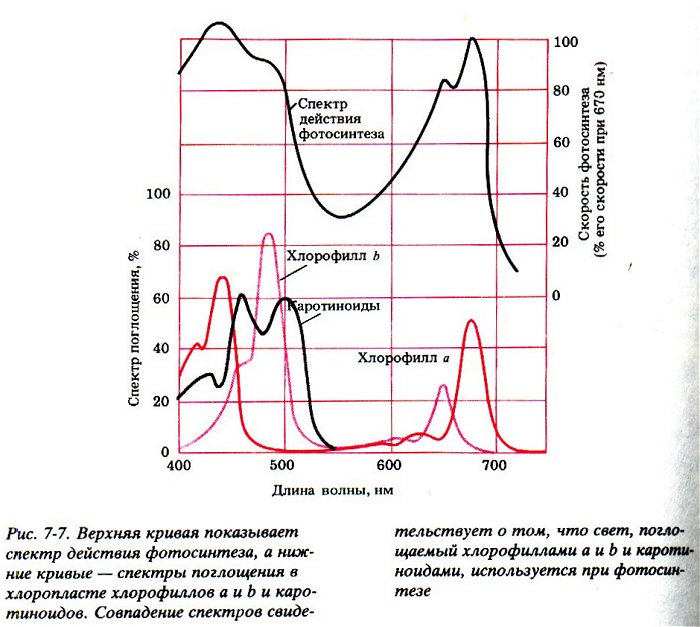
Молекула хлорофилла поглощает фотоны в фиолетовой и синей, а затем в красной части спектра, и не взаимодействует с фотонами в зеленой и желтой части спектра.
Поэтому хлорофилл и растения выглядят зелеными – они попросту никак не могут воспользоваться зелеными лучами и оставляют их гулять по белу свету (делая его тем самым зеленее).
Пигменты фотосинтеза располагаются на внутренней стороне мембраны тилакоидов.
Пигменты организованы в фотосистемы (антенные поля по улавливанию света) – содержащие по 250–400 молекул разных пигментов.
Фотосистема состоит из:
реакционного центра фотосистемы (молекула хлорофилла а),
антенных молекул
Все пигменты в фотосистеме способны передавать друг другу энергию возбужденного состояния. Энергия фотона, поглощенная той или иной молекулой пигмента, переносится на соседнюю молекулу, пока не достигнет реакционного центра. Когда резонансная система реакционного центра переходит в возбужденное состояние, она передает два возбужденных электрона молекуле-акцептору и тем самым окисляется и приобретает положительный заряд.
У растений:
фотосистема 1 (максимум поглощения света на длине волны 700 нм - Р700)
фотосистема 2 (максимум поглощения света на длине волны 680 нм - Р680
Различия в оптимумах поглощения обусловлены небольшими различиями в структуре пигментов.
Две системы работают сопряженно, как конвейер, состоящий из двух частей и называющийся нециклическим фотофосфорилированием.
Суммарное уравнение для нециклического фотофосфорилирования:
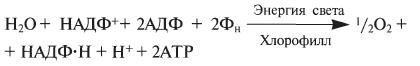
Ф - условное обозначение остатка фосфорной кислоты
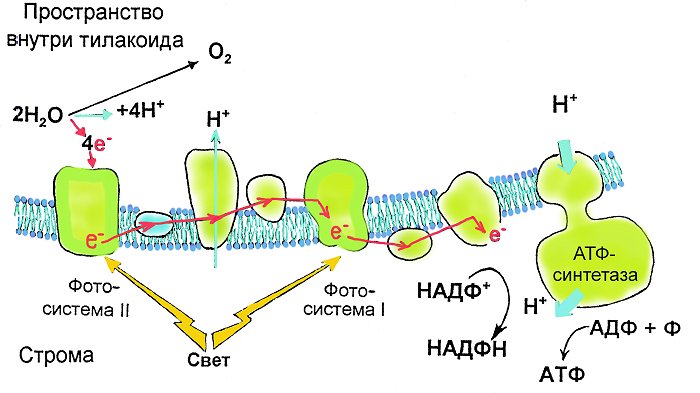
Цикл начинается с фотосистемы 2.
1) антенные молекулы улавливают фотон и передают возбуждение молекуле активного центра Р680;
2) возбужденная молекула Р680 отдает два электрона кофактору Q при этом она окисляется и приобретает положительный заряд;
Кофактор (cofactor). Кофермент или любое другое вещество, необходимое для выполнения ферментом его функции
Коферменты (коэнзимы) [от лат. co (cum) — вместе и ферменты], органические соединения небелковой природы, участвующие в ферментативной реакции в качестве акцепторов отдельных атомов или атомных групп, отщепляемых ферментом от молекулы субстрата, т.е. для осуществления каталитического действия ферментов. Эти веществава, в отличие от белкового компонента фермента (апофермента), имеют сравнительно небольшую молекулярную массу и, как правило, термостабильны. Иногда под Коферментами подразумевают любые низкомолекулярные вещества, участие которых необходимо для проявления каталитического действия фермента, в т. ч. и ионы, напр. К+, Mg2+ и Мn2+ . Располагаются оферменты. в активном центре фермента и вместе с субстратом и функциональными группами активного центра образуют активированный комплекс.
Для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Исключение составляют гидролитические ферменты (например, протеазы, липазы, рибонуклеаза), выполняющие свою функцию в отсутствие кофермента.
Молекула восстанавливается Р680 (под действием ферментов),. При этом вода диссоциирует на протоны и молекулярный кислород, т.е. вода является донором электронов, который обеспечивает восполнение электронов в Р680.
ФОТОЛИЗ ВОДЫ — расщепление молекулы воды, в частности в процессе фотосинтеза. Вследствие фотолиза воды образуется кислород, выделяющийся зелеными растениями на свету.
Общий ход фотолиза воды.
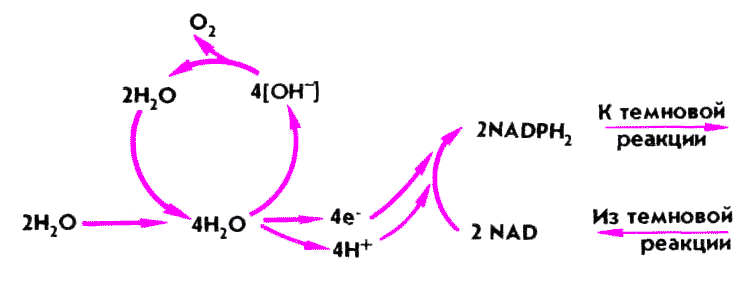
Уравнение фотолиза воды
hv
2Н20=4Н+ 4е- +02
Для создания одной молекулы кислорода нужно восстановить две молекулы Р680, потерявшие в сумме четыре электрона, при этом образуются четыре протона.
4) эти протоны образуются во внутреннем пространстве тилакоида, где создается избыточная концентрация протонов по сравнению с окружающим пространством (т. е. более кислая среда), (формируются - протонный градиент и мембранный потенциал).
АТФ-синтетаза парами выпускет протоны наружу и синтезирует АТФ из АДФ.
5) два электрона, поступившие к кофактору Q, передаются далее по цепочке белков, которая очень похожа на цепь переноса электронов.
6) электроны поступают от пластоцианина к реакционному центру фотосистемы 1 – молекуле Р700.
Фотосистема 1:
1) реакционный центр Р700 отдает 2e акцепторному железосодержащему белку (Р430), окисляется и приобретает положительный заряд;
2) эта молекула восстанавливается и теряет заряд, получив два «успокоившихся» (но не до исходного состояния – их энергия еще не до конца израсходована!) электрона, изначально поступивших от фотосистемы 2. В этом случае необходимости в фотолизе нет и его не происходит;
3) Р430 отдает электроны белку ферродоксину;
4) получив электроны, этот белок восстанавливает кофермент НАДФ+ до НАДФ∙Н. Данный кофермент представляет собой фосфорилированный НАД. Процесс происходит на внешней мембране тилакоида.
Таким образом, по цепи белков передаются электроны, в результате чего во внутреннем пространстве тилакоида создается избыточная концентрация протонов, а на мембране – разность потенциалов.
Возникающая потенциальная энергия используется для синтеза АТФ за счет движения протонов по градиенту, осуществляемого АТФ-синтетазой. (т.о. энергия потока используется для получения АТФ)
Для возбуждения электронов используется свет
НАДФ + используется в темновой стадии фотосинтеза (а может быть далее использован и митохондриях).
В целом получается, что протоны образуются во внутреннем пространстве тилакоида при фотолизе воды, закачиваются туда же в ходе работы фотосистемы 2 и черпаются из внешнего пространства тилакоида для восстановления НАДФ+ до НАДФ-Н (т.е. электроны, движущиеся вниз по градиенту энергии к НАДФ вдоль цепи переноса электронов, взаимодействуют с ионами водорода (из воды), образуя восстановленный НАДФ-)
Вот на схеме более или менее показаны все основные процессы световой стадии фотосинтеза:
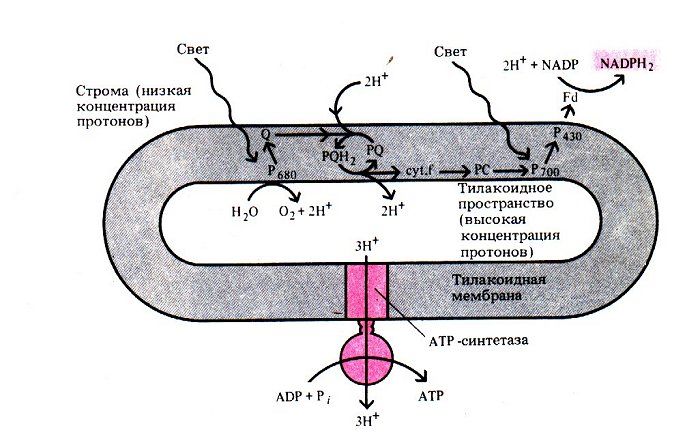
Фотосистема 1 может работать автономно.
Циклическое фотофосфорилирование
При этом используется обходной путь переноса электронов от возбужденного реакционного центра – а именно та же цепь переноса электронов, которая ведет из фотосистемы 2. Электроны проходят по ней возвращаются обратно к реакционному центру фотосистемы 1 – Р700. Т.о. электроны возвращаются обратно к Р700 по другой цепи переноса электронов. Как и при нециклическом фосфорилировании энергия возбуждения электронов, перемещающихся вдоль этой цепи, направляется на получение АТФ.
Оно может идти параллельно с нециклическим. Кроме того, оно используется некоторыми фотосинтетическими бактериями, которые в процессе фотосинтеза не выделяют кислорода.
Сравнение циклического и нециклического фотофосфорилирования

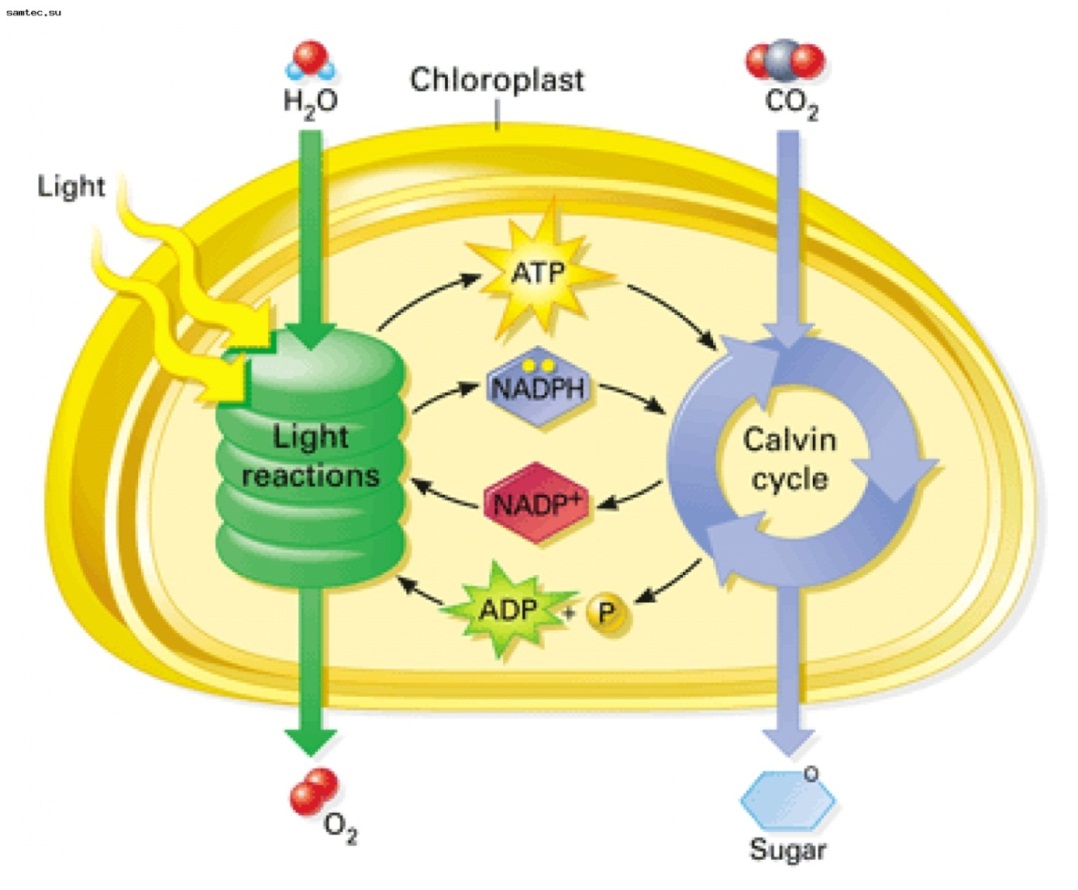
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами:
1) синтезом АТФ;
2) образованием НАДФ·Н2;
3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
количество АТФ, синтезируемое АТФ-синтетазой, не связано жесткой зависимостью от количества электронов, пропущенных по цепочке белков в фотосистеме II.
Темновая фаза
Протекает в строме хлоропласта.
Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте.
Реакции темновой фазы представляют цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
В настоящее время различают два типа фотосинтеза: С3- и С4-фотосинтез. Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин).
Характерные особенности С3-фотосинтеза:
1) Первая реакция в этой цепочке - фиксация углекислого газа. Акцептором углекислого газа является рибулозодифосфат,
2) реакцию карбоксилирования рибулозодифосфата катализирует рибулозодифосфат-карбоксилаза,
3) в результате карбоксилирования рибулозодифосфат образуется шестиуглеродное соединение, которое распадается на две фосфоглицериновой кислоты.
фосфоглицериновая кислота восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию рибулозодифосфата, часть превращается в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу;
цикл этих реакций получил название цикл Кальвина:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды.
С3-фотосинтез
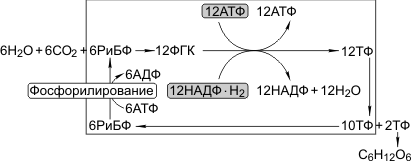
Путь углерода в фотосинтезе (цикл Кальвина)
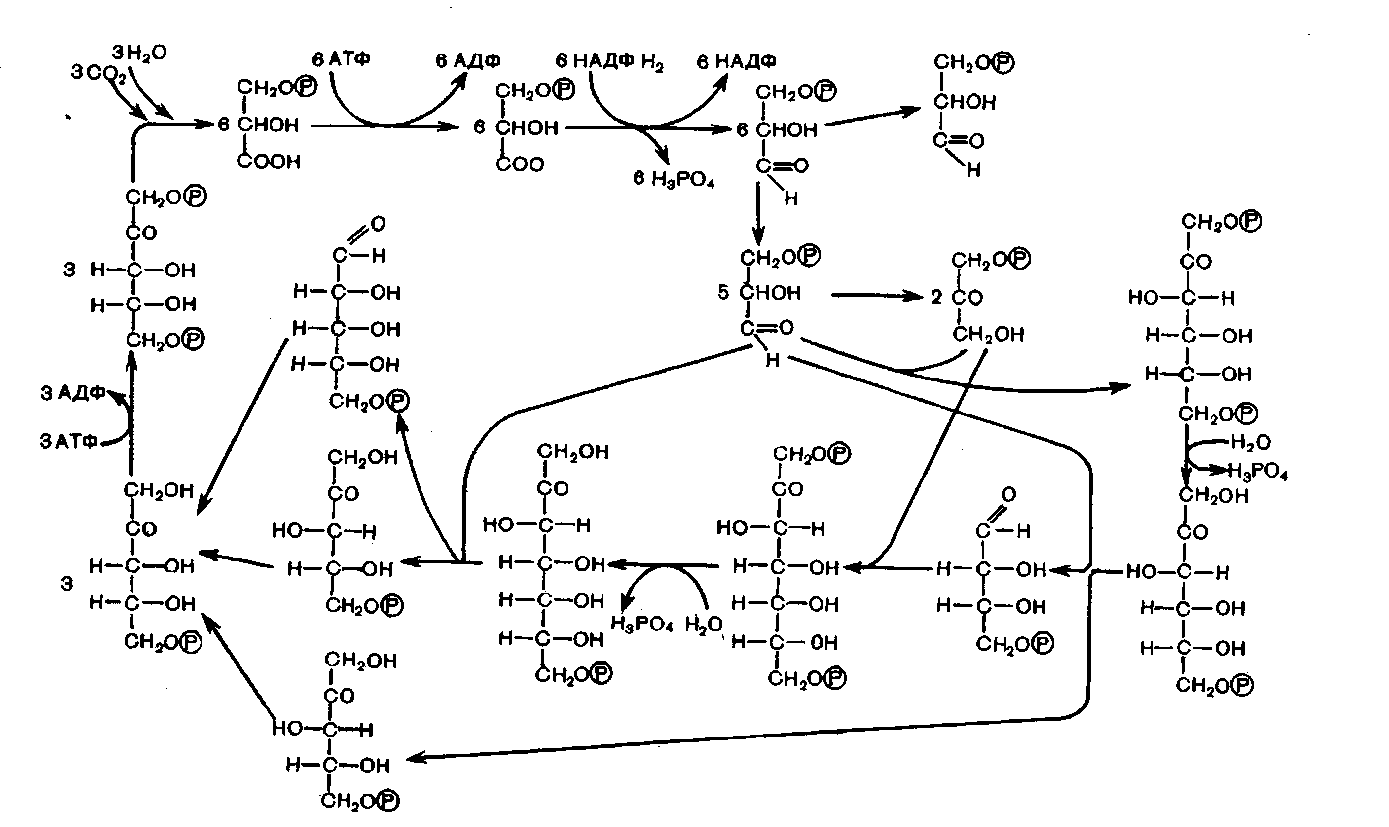
Схема фотосинтеза
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
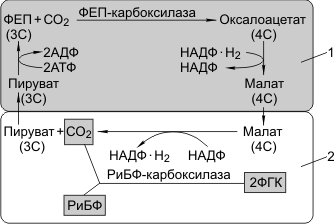
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
С4-фотосинтез:
1 — клетка мезофилла; 2 — клетка обкладки проводящего пучка.
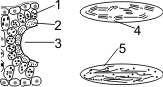
Строение С4-растений:
1 — наружный слой — клетки мезофилла; 2 — внут�ренний слой — клетки обкладки; 3 — «Кранц-анатомия»; 4, 5 — хлоро�пласты; 4 — много�числен�ные граны, крахмала мало; 5 — немного�числен�ные граны, крахмала много.
|
Фотодыхание
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для рибулозодифосфат - карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиДФ → фосфогликолат (2С) + ФГК (3С).
ФГК - фосфоглицериновая кислота
Фермент при этом называется РиДФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2.
Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
Пероксисомы клеточные органеллы диаметром
0,5 мкм, имеющиеся во всех эукариотических клетках и состоящие из одинарной мембраны происходящей из из гладкого эндоплазматического ретикулума. Не содержат ДНК; все белки приходят из цитозоля.
Содержат окислительные ферменты (пероксидаза, каталаза, уратоксидаза, оксидаза-D-аминокислот) в высокой концентрации, Наряду с митохондриями являются главным местом утилизации O2. В пероксисомах происходит окисление жирных кислот, с образованием ацетил-CoA, У млекопитающих окисление жирных кислот происходит в митохондриях и пероксисомах, у дрожжей и растений - только в пероксисомах.
Фотодыхание
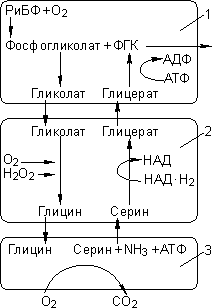
Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом.
К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ.
АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Бактериальный фотосинтез
Фотосинтез бактериальный — фотосинтез, осуществляемый бактериальными микроорганизмами.
Типично водные микроорганизмы, распространенные в пресных и соленых водоемах. Особенно часто они встречаются в местах, где есть сероводород, как в мелководье, так и на значительной глубине. В почве фототрофных бактерий мало, но при затоплении ее водой они могут расти весьма интенсивно.
Пурпурные и зеленые бактерии — наиболее древние фотосинтезирующие организмы
Всего сейчас описано около пятидесяти видов пурпурных и зеленых бактерий.
Содержат развитую систему мембран, некоторые из них выполняют функцию фотосинтезирующего аппарата (хроматофоры или тилакоидами)
В отличие от растений, фотосинтез у пурпурных и зеленых бактерий
не сопровождается выделением кислорода
Донорами водорода при ассимиляции углекислоты служит не вода, а другие вещества:
сероводород (2C02+H2S+2H20 -> 2(CH20)+H2S04),
молекулярная сера
тиосульфат
сульфит
молекулярный водород
органические соединения.
Обязательными компонентами сред, кроме источников углерода и азота, естественно, являются фосфор, сера, калий и магний в виде минеральных солей. Установлено также, что эти микроорганизмы нуждаются в довольно большом количестве железа, но по сравнению с растениями проявляют значительно меньшую потребность в марганце. Рост их зависит от концентрации кальция и наличия в небольших количествах таких элементов, как Mo, Co, Zn, Си и, видимо, других.
У многих пурпурных и зеленых бактерий установлена способность фиксировать молекулярный азот.
Наличие максимумов поглощения бактериохлорофиллов в области 800—1100 нм обеспечивает возможность роста фототрофных бактерий, особенно пурпурных, при наличии только невидимых инфракрасных лучей.
Различия в спектрах поглощения отдельных представителей фототрофных бактерий, а также фототрофных бактерий и растений имеют экологическое значение, позволяя развиваться им в одних и тех же местах.
Кроме хлорофиллов, все фототрофные бактерии содержат каротиноиды, состав которых у разных видов неодинаков и достаточно разнообразен. Всего к настоящему времени у фототрофных бактерий обнаружено около шестидесяти каротиноидов,
Размножение клеток
Размножение или пролиферация (от лат. proles — потомство, ferre — нести) клеток — это процесс, который приводит к росту и обновлению клеток. Данный процесс характерен как для одноклеточных, так и многоклеточных организмов.
Соматические клетки многоклеточных организмов размножаются путем митотического деления
Митоз: способ упорядоченного деления клеток, при котором каждая из двух дочерних клеток получает хромосомы в количестве и имеющие строение как у материнской клетке. При каждом митозе образуется копия каждой хромосомы и действует точный механизм их распределения между дочерними клетками.
В митотическом делении клетки различают две стороны:
кариокинез (разделение исходного ядра на два дочерних)
цитокинез (разделение цитоплазмы с образованием двух дочерних клеток)
Кариокинез и цитокинез протекают синхронно
Митотический цикл - совокупность процессов, происходящих в клетке от одного деления до другого. Состоит из двух стадий:
интерфаза (стадии покоя)
митоз (стадии деления)
Термины «митоз» и «кариокинез» — синонимы
Интерфаза
предшествует митозу, в ней происходит синтез ДНК (длительность составляет не менее 90% клеточного цикла)
Различают три периода интерфазы;
пресинтетический (G1)
синтетический (S) -
постсинтетический (G2)
1 12-24 часа
Синтез ДНК (около 5 часов)
Около 4 часов
Митоз (около 1,5 часов)
|
Митоз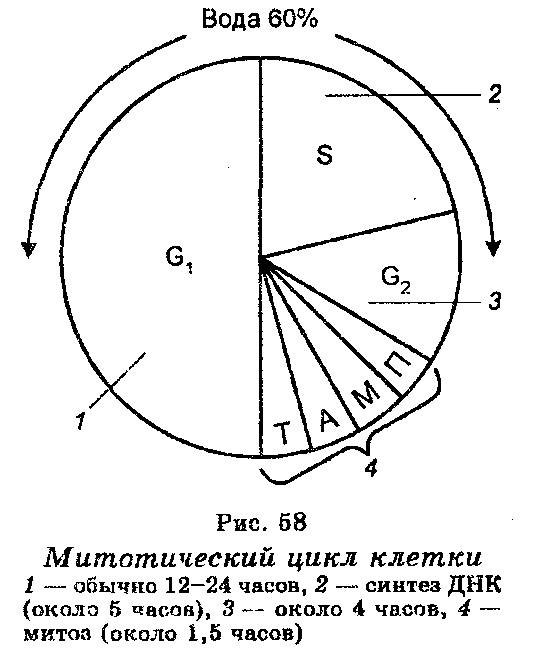
Состоит из четырех фаз:
профаза
метафаза
анафазы
телофазы
Митотический цикл и митоз
Фазы
|
Процесс, происходящий в клетке
|
(фаза ме�жду делени�ями клеток)
|
Пресинтетический период (G1)
|
Происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК. Увеличивается количество митохондрий. Обычно этот период длится 12-24 часа.
|
Синтетический период (S)
|
Синтез (репликация) ДНК, в результате чего количество ее удваивается; синтез РНК и белков. Т.о, наиболее фундаментальной особенностью S-периода является репликация генов и удвоение набора генов каждой дуплицированной хромосомы (двухроматидные хромосомы).
Длительность S-периода обычно составляет около 5 часов.
|
Постсиитетический период (G2)
|
Остановка синтеза ДНК и накопление энергии; продолжается синтез РНК и белков, формирующий нити веретена деления. Длительность составляет 3—6 часов.
|
Ми�тоз
|
Профаза
(пер�вая фаза деления)
|
Двухроматидные хромосомы спирализуются, ядрышки растворяют�ся, центриоли расходятся, ядерная оболочка растворяется, образуют�ся нити веретена деления (построено из микротрубочек и различных белков). Длительность составляет примерно 30-60 минут.
|
Метафаза
(фа�за скопления хро�мосом)
|
Нити веретена деления присоединяются к центромерам хромо�сом, двухроматидные хромосомы сосредоточиваются на экваторе клетки. Длительность метафазы составляет 2—10 минут
|
Анафаза
(фа�за расхождения хромосом)
|
Центромеры делятся, однохроматидные хромосомы растягиваются нитями веретена деления к полю�сам клетки. Анафаза длится 2-3 минуты
|
|
Телофаза
(фа�за окончания де�ления)
|
Однохроматидные хромосомы деспирализуются, сформировывает�ся ядрышко, восстанавливается ядерная оболочка, на экваторе на�чинает закладываться перегородка между клетками, растворяются нити веретена деления
Образуется ядерная оболочка, вновь появляется ядрышко. Длительность составляет 20-30 минут.
|
Продолжительность митотических циклов разных клеток различна и составляет от нескольких часов до нескольких дней. Однако она зависит от типа тканей, физиологического состояния, внешних факторов (температура, свет).
Разные ткани характеризуются разной митотической активностью.
В зависимости от митотической активности различают ткани:
стабильные (клетки не делятся, количество клеточной ДНК постоянно)
Пример: клетки центральной и периферической нервной системы. В этих клетках происходят лишь возрастные изменения
растущие (клетки живут всю жизнь, но среди последних имеются такие, которые делятся посредством митоза). Приводит к увеличению размеров органов.
Пример: ткани почек, желез внутренней секреции, скелетная и сердечная мускулатуры
обновляющиеся (многие клетки подвержены митозам, в результате чего погибающие клетки компенсируются вновь образующимися)
Пример: клетки желудочно-кишечного, дыхательного и мочеполового трактов, эпидермиса, костного мозга, семенников
У высших организмов митотическое деление клеток обеспечивает их рост с последующим увеличением массы тела и дифференциацией клеток.
Для деления клеток млекопитающих и птиц характерно то, что оно имеет определенные ограничения количества клеточных удвоений.
Пример: фибробласты плодов человека удваиваются лишь на протяжении 50 генераций, тогда как фибробласты от людей в возрасте 40 и 80 лет подвергаются примерно 40 и 30 удвоениям соответственно, если их культивируют в стандартных условиях (явление старения клеток).
В организме большинство клеток стареет:
клетки печени живут около 18 месяцев
эритроциты — 4 месяца, в результате чего в них накапливаются липиды, кальций, пигмент «изнашивания» и они гибнут.
Подсчитано, что организм взрослого человека ежедневно теряет около 1-2% своих клеток в результате их гибели.
После смерти клетки в ней происходит коагуляция протоплазмы, распад митохондрий и других органелл в результате аутолиза (активации внутриклеточных ферментов).
Считается, что клеточное содержание организма человека обновляется примерно каждые семь лет.
Особенно сильно замещение клеток происходит в крови
Эпителий тонкого кишечника человека — каждые 7-8 дней.
Нервные клетки функционируют (живут) на протяжении всей жизни организмов.
Амитоз
Амитоз - прямое деление ядра клетки.
При амитозе сохраняется интерфазное состояние ядра, ядрышко, ядерная мембрана.
Ядро клетки делится на две части без формирования веретена, в результате чего образуется двухъядерная клетка.
Амитоз - аномальный механизм в размножении клеток (встречается иногда в клетках скелетной мускулатуры, кожного эпителия, соединительной ткани).
Мейоз
Происходит при образовании гамет (сперматозоидов и яйцеклеток).
Исходная клетка имеет диплоидный набор хромосом, которые затем удваиваются.
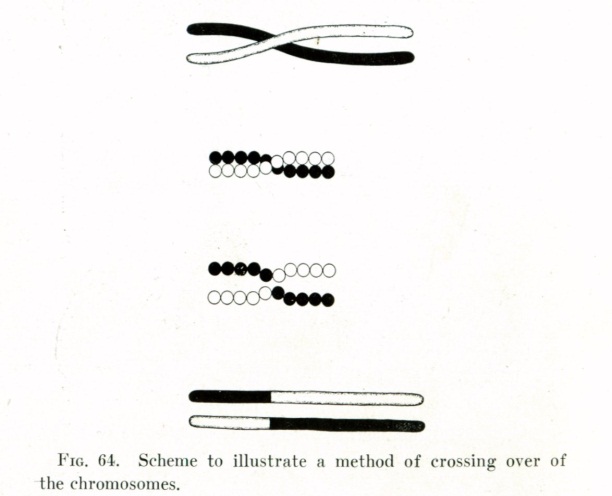
|
При мейозе происходит
кроссинговер - обмен гомологичными участками хромосом.
|
Первое деление мейоза
Новые хромосомы расходятся и образуются клетки с диплоидным набором хромосом, но состав этих хромосом отличается от исходного - в них произошла рекомбинация.
Рекомбинация - перераспределение (перекомбинирование) генетического материала родителей, в результате чего у потомков появляются новые сочетания генов, определяющие новые сочетания признаков.
Это основа комбинативной изменчивости. У эукариотических организмов, размножающихся половым путём, рекомбинация происходит в мейозе при независимом расхождении хромосом и при обмене гомологичными участками между гомологичными хромосомами (кроссинговере). Возможна и т. н. незаконная рекомбинация, когда структурные перестройки затрагивают негомологичные хромосомы. Рекомбинации бывают и в половых, и, гораздо реже, в соматических клетках.
Напоминание: при митозе в каждой хромосоме
хроматиды просто расходятся
Второе деление мейоза
Фазы
|
Процесс, происходящий в клетке
|
Первое (I) мейотическое деление
|
профаза I
|
Сближение гомологичных хромосом (точка каждой хроматиды одной хромосомы совме�щается с соответствующей точкой хроматиды другой, гомологичной хромосомы)
Процесс точного и тесного сближения гомо�логичных хромосом в мейозе называют конъюгацией.
Кроссинговер обмен участками гомологичных хромосом в процессе клеточного деления, что приводит к новому сочетанию генов и к изменению фенотипа.
|
метафаза I
|
Спирализация хромосом максимальна
Конъюгированные хромосомы располагаются по экватору. К ним прикрепляются нити веретена деления.
|
анафаза I
|
хромосомы расходятся к различным полюсам.
хромосомный набор гаплоиден
каждая хромосома состоит из двух хроматид (1n2с)
|
телофаза I
|
восстанавливается ядерная оболочка
ма�теринская клетка делится на две дочерние
|
Второе (II) мейотическое деление.
|
Протекает так же, как обычное митотическое деление, с той лишь разницей, что делящаяся клетка гаплоидна (1n2с).
|
профаза II
|
по периферии ядра располагаются нитевидные хромосомы - униваленты, образуется веретено деления, хромосомы, приближаются к плоскости экватора
|
метафаза II
|
хромосомы выстраиваются вдоль экватора. К ним подходят нити веретена деления
|
|
анафаза II
|
хроматиды расходятся и увлекаются нитями веретена от плоскости экватора к противоположным полюсам.
|
|
телофаза II
|
хромосомы истончаются, образуя нити, и у полюсов формируются ядра дочерних клеток.
|
происходит без синтеза ДНК, поэтому при этом делении количество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором.
Из одной диплоидной клетки образуются четыре гаплоидных клетки.
В каждом деление мейоза выделяют четыре стадии:
профаза
метафаза
анафаза
телофаза
Красным обозначены материнские хромосомы, синим - отцовские.
Клетки-организмы (одноклеточные организмы) размножаются простым делением надвое (бактерии, саркодовые), множественным делением (споровики и др.) или другим путем. Поэтому у бактерий и одноклеточных животных удвоение клеток представляет собой размножение их как самостоятельных организмов, поскольку из исходной формы (организма) образуется две новые клетки, каждая из которых является организмом. Каждая дочерняя клетка (организм) получает полную
генетическую информацию, несомую исходной клеткой-организмом.
мейоз
Биологическое значение мейоза:
образуются хромосомы обновленного генетического состава благодаря кроссинговеру между гомологичными хромосомами;
достигается наследственная разнородность гамет, так как во время первого мейотического деления из пары гомологичных хромосом в одну из двух гамет отходит материнская хромосома, в другую - отцовская;
после оплодотворения гаплоидные гаметы (1n1с) от отца и матери создают диплоидное ядро зиготы с числом хромосом, присущим данному виду.
мейоз — основа комбинативной генетической изменчивости
Вопросы для самоконтроля
Какие элементы относятся к макроэлементам, какие к микроэлементам?
Привести примеры роли микроэлементов в клетке?
Назовите мономеры белков, жиров, углеводов, нуклеиновых кислот.
Перечислите функции белков, жиров, углеводов, нуклеиновых кислот.
Перечислите органоиды и органеллы клетки
В чем отличие растительной и животной клетки?
Что такое анаболизм и катаболизм?
Назовите основные пути поступления веществ в клетки
В чем суть процесса фотосинтеза?
Что такое фотодыхание?
Что такое хемосинтез?
Перечислите основные стадии митоза.
В чем особенность мейоза, его биологически смысл?
|
 Скачать 5.67 Mb.
Скачать 5.67 Mb.