методичка БРР. Учебнометодическое пособие для студентов очной и очнозаочной формы обучения по специальности 020201 биология
 Скачать 19.97 Mb. Скачать 19.97 Mb.
|

|
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ государственное образовательное учреждение высшего профессионального образования «Нижегородский государственный университет им. Н. И. Лобачевского» Биологический факультет Кафедра зоологии «БИОЛОГИЯ РАЗМНОЖЕНИЯ И РАЗВИТИЯ». УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ ДЛЯ СТУДЕНТОВ ОЧНОЙ И ОЧНО-ЗАОЧНОЙ ФОРМЫ ОБУЧЕНИЯ по специальности 020201 - БИОЛОГИЯ Нижний Новгород 2008 УДК 591.3 ББК Е 63 Ф–15 «Биология размножения и развития». Учебно-методическое пособие для студентов очной и очно-заочной форм обучения / Сост. Г.А. Фадеева, У.В. Колова, С. А. Мельник. – Н. Новгород: Нижегородский государственный университет, 2008. – 69 с. Пособие содержит теоретический материал для подготовки к занятиям по биологии размножения и развития, а также рисунки и описание препаратов, рассматриваемых на занятиях. Составители: к. б. н. доц. Г. А. Фадеева, к. б. н., ст. преп. У. В. Колова, к. б. н., ст. преп. С. А. Мельник Рецензенты: докт. биол. наук, профессор Охапкин А.Г. © Нижегородский государственный университет им. Н.И. Лобачевского, 2008 Введение Биология размножения и развития изучает развитие многоклеточных животных. В основу ее положены эмбриологические исследования, которые развиваются в последние годы с использованием открытий в областях, смежных с эмбриологией, а именно, с цитологией, генетикой, молекулярной биологией и др. Благодаря успехам молекулярной биологии и генетики, в эмбриологии решаются такие фундаментальные проблемы, как функции генов в развитии и регуляции генной экспрессии, индукционные взаимодействия, проблемы регенерации и др. Знание закономерностей регуляции онтогенеза и условий его изменения необходимо для понимания механизмов эволюции и управления онтогенезом. ГАМЕТОГЕНЕЗ Клетки организма подразделяются на соматические и половые. У большинства организмов (насекомые, позвоночные и др.) обособление половых клеток происходит на ранних этапах эмбрионального развития, в основе этого процесса лежит ооплазматическая сегрегация яйцеклетки. В отличие от соматических клеток, половые клетки тотипотентны, т.е. они сохраняют способность образовывать органы и ткани. Половые клетки у всех животных, имеющих морфологически выраженные гонады, закладываются вне гонады (экстрагонадно). С момента вселения в гонаду их называют первичными половыми клетками. У большинства животных первичные половые клетки – единственный источник зрелых гамет, они активно мигрируют посредством амебоидного движения к зачаткам гонад, преодолевая клеточные пласты. Локализация первичных половых клеток в гонадах различна. Так, в случае сперматогенеза первичные половые клетки пролиферируют через ткани и локализуются в периферической части семенных канальцев. При оогенезе первичные половые клетки локализуются в корковом слое гонады и окружаются фолликулярными клетками. Занятие 1 Сперматогенез Сперматогенез – это процесс образования мужских гаплоидных клеток (гамет). Сперматогенез состоит из следующих этапов:
В течение периода размножения диплоидные клетки – сперматогонии – делятся путем митоза несколько раз. У некоторых животных, например губок и кишечнополостных, истинные гонады отсутствуют, поэтому сперматогенез происходит в рассеянных по телу сперматогенных скоплениях. Число митотических делений у каждого вида строго специфично и варьирует от 3 до 5. Крайние варианты отмечены для клещей (2 деления) и рыб (до 14 делений). Последующие генерации сперматогониев отличаются по степени конденсации хроматина и величине. У делящихся сперматогониев процесс цитотомии завершается не полностью. Все клетки, образующиеся из одного сперматогония, образуют единый клеточный клон, а между ними формируются цитоплазматические мостики (фузомы). Как и всякая обновляющаяся ткань, популяция мужских половых клеток содержит и стволовые клетки. Эти клетки расположены изолированно, у них понижена митотическая активность. Второй период сперматогенеза – рост. Половые клетки на этой стадии называются сперматоцитами первого порядка. Они несколько увеличиваются в объеме, а в ядрах проходит профаза первого мейотического деления. В этот период осуществляется процесс конъюгации хромосом, которые скрепляются друг с другом посредством образования синаптонемального комплекса. Он сползает с гомологичных хромосом на стадии диплотены. В течение третьего периода – созревания, совершаются два деления и редукция числа хромосом. После первого деления созревания клетка сперматогенного ряда называется сперматоцитом второго порядка, а после второго – сперматидой. Четвертый завершающий период – период формирования или спермиогенез, когда сперматида после ряда преобразований превращается в сперматозоид. Сперматозоид состоит из головки, шейки и жгутика (в единичных случаях жгутик может отсутствовать). В процессе формирования значительно уменьшается объем ядра за счет конденсации хроматина и элиминации кариоплазмы, при этом хроматин превращается в биологически инертную массу. Ядро смещается в ту часть клетки, которая впоследствии становится головкой сперматозоида. Формируется специфическая для сперматозоидов органелла – акросома. Она образуется из аппарата Гольджи и располагается в переднем отделе головки. Чаще всего акросома имеет форму колпачка или пузырька, содержит лизирующие ферменты. Центриоли перемещаются под ядро. Дистальная центриоль становится базальным тельцем, являющимся основанием для жгутика. Жгутик сперматозоида имеет типичное строение и обозначается формулой 9 (2) + 2. Митохондрии также перемещаются под ядро и окружают кольцом или спиралью основание жгутика. Митохондрии вырабатывают энергию, необходимую для движения сперматозоида. Сперматиды утрачивают во время спермиогенеза большую часть цитоплазмы и некоторые др. органеллы, которые резорбируются клетками Сертоли. К концу периода формирования происходит индивидуализация сперматозоидов и приобретение ими видоспецифичных признаков. В эякуляте сперматозоиды суспензированы в семенной жидкости, которая является смесью секретов желез мужского репродуктивного тракта. Значения относительной концентрации сперматозоидов в эякуляте очень важны для успешного оплодотворения и колеблются от 50 до 170 млн. в 1 мл. Препарат 1. Семенник крысы (гистологический срез) На срезе семенника канальцы выглядят либо округлыми, либо овальными – это зависит от плоскости сечения. Между семенными канальцами расположена соединительная ткань и клетки Лейдига, основная функция которых – выработка мужского полового гормона тестостерона, необходимого для осуществления сперматогенеза. Стенку канальца образуют несколько слоев клеток, которые представляют собой дифференцирующиеся половые клетки (рис. 1). Развитие их происходит в направлении от базальной мембраны в просвет канальца. Помимо сперматогенного эпителия (сперматогониев и их потомков) в состав стенки канальца входят соматические по происхождению клетки Сертоли. Основания этих клеток находятся на стенке канальца, а разветвленные их части простираются внутрь канальца. Рассмотреть границы клеток Сертоли невозможно, но в основании каждой клетки располагаются ядра треугольной формы. Функции клеток Сертоли различны: трофическая, опорная, регуляторная, фагоцитарная. Между основаниями этих клеток располагаются сперматогонии. Они характеризуются относительно небольшими размерами и крупным ядром. Хроматин в ядрах сперматогониев первой генерации деконденсирован, равномерно распределен в кариоплазме. Ярус семенного канальца (базальный компартмент), в котором расположены сперматогонии, обозначается зоной размножения. В околополостном отсеке ярусами располагаются последовательно сперматоциты первого порядка, спематоциты второго порядка и сперматиды. При формировании спермотозоидов от сперматид отделяются резидуальные тельца. В самом центре канальцев располагаются сперматозоиды. На рисунке 2 показана схема строения того же семенного канальца, позволяющая детально разобраться в строении сперматогенного эпителия. 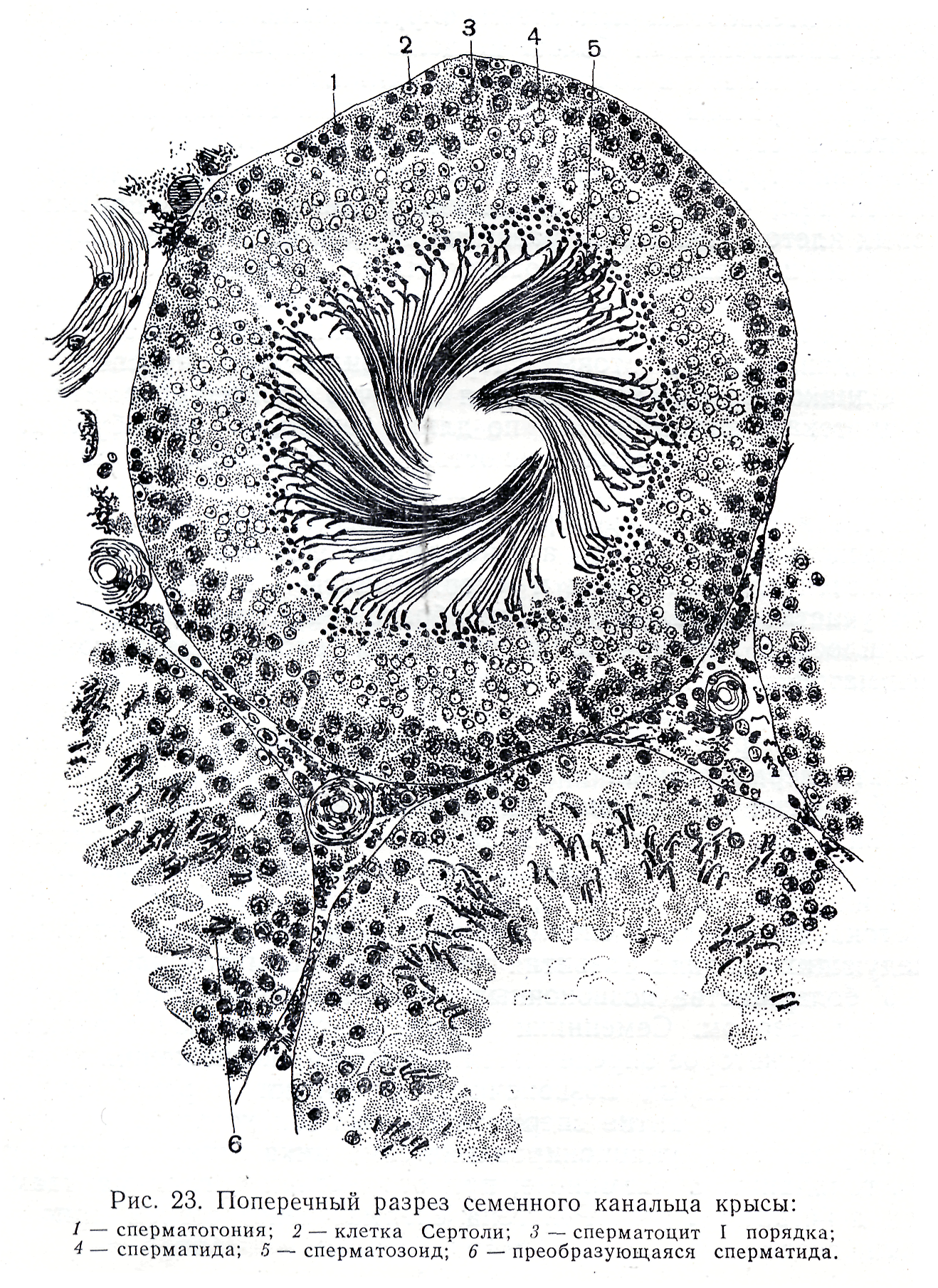 Рис. 1. Поперечный разрез семенного канальца крысы (Мануилова, 1964). 1 – сперматогония; 2 – клетка Сертоли; 3 – сперматоцит первого порядка; 4 – сперматида; 5 – сперматозоид; 6 – преобразующаяся сперматида. 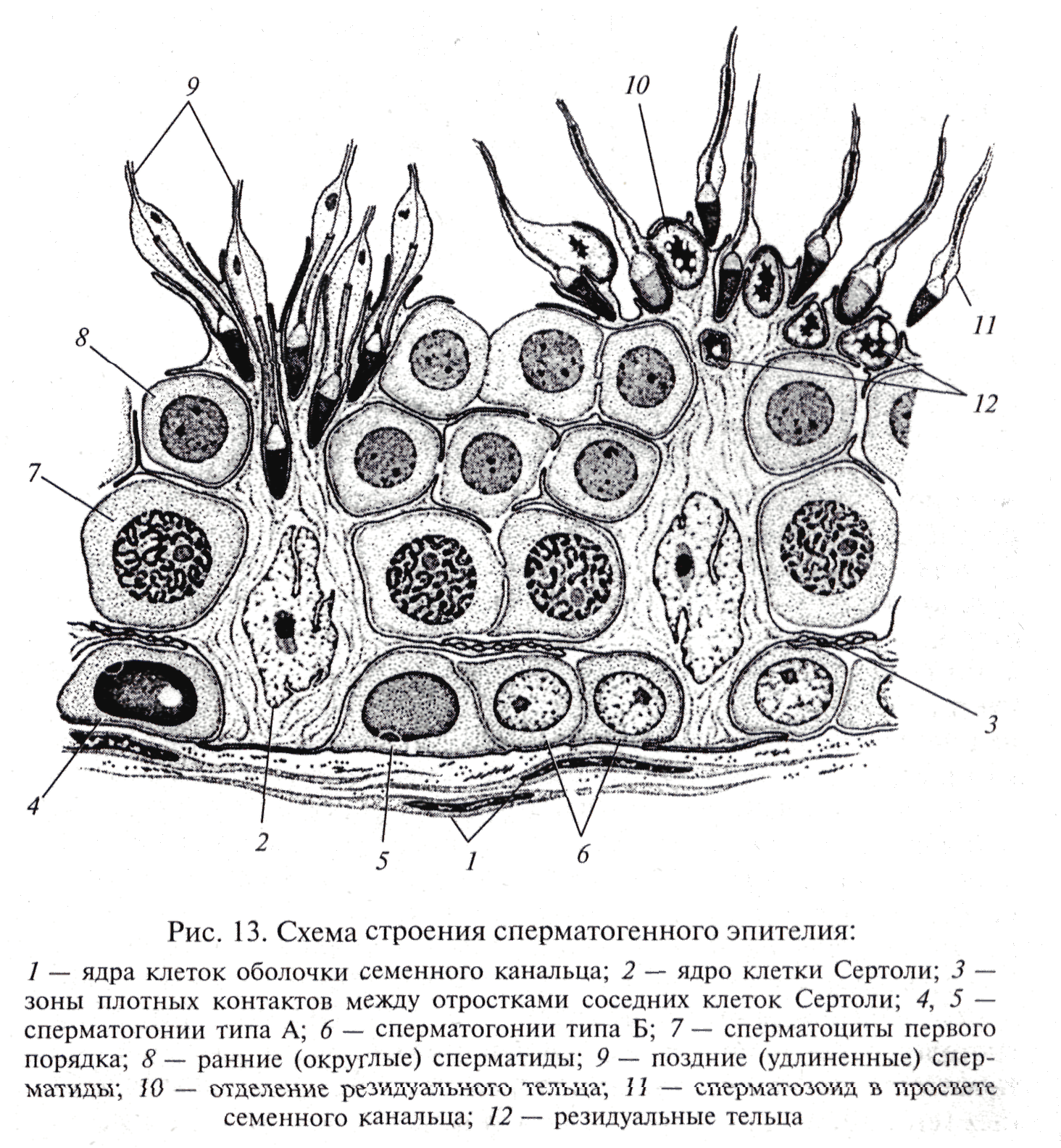 Рис. 2. Схема строения сперматогенного эпителия (Мануилова, 1964). 1 – ядра клеток оболочки семенного канальца; 2 – ядро клетки Сертоли; 3 – зоны плотных контактов между отростками соседних клеток Сертоли; 4,5,6 – сперматогонии; 7 – сперматоциты первого порядка; 8 – ранние (округлые) сперматиды; 9 – поздние (удлиненные) сперматиды; 10 – отделение резидуального тельца; 11 – сперматозоид в просвете семенного канальца; 12 – резидуальные тельца. Препарат 2. Сперматозоиды морской свинки Сперматозоиды морской свинки – это типичные жгутиковые клетки. Общая длина их равна 100 мкм. Отчетливо выражена головка, шейка и хвостик. Головка имеет округлую форму, центр ее занимает ядро. Акросома темная и в виде шапочки расположена на апикальном конце клетки. Область шейки несколько утолщена. Здесь сохраняется резидуальная цитоплазма в виде капельки. Капелька обычно исчезает по мере прохождения сперматозоида по канальцам придатка семенника. Хвостик длинный, покрыт цитоплазматической оболочкой, состоит их осевой нити и окружающей ее цитоплазмы. Иногда на препаратах можно увидеть «многохвостые» спермии. Это результат склеивания нескольких спермиев, которые обладают высокой способностью к агглютинации 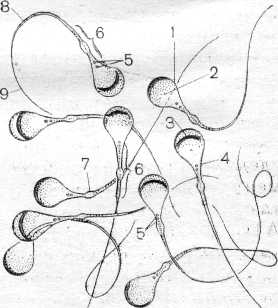 . Рис. 3. Сперматозоиды морской свинки (Новиков, Святенко, 1984). 1 – головка; 2 – ядро; 3 – акросома; 4 – шейка; 5 – две центриоли; 6 – связующий отдел; 7 – утолщение; 8 – главный отдел хвостика; 9 – концевой отдел хвостика. Препарат 3. Сперматозоиды тритона Сперматозоиды тритона, как и морской свинки, относятся к жгутиковым формам. Размер клетки примерно равен 500 мкм. Головка сильно вытянута и заострена, в апикальной части имеет форму гарпуна. Под акросомой располагается крупное плотное стержневидное ядро. К задней его части примыкает очень короткая шейка. Цитоплазматическая оболочка длинного хвоста формирует ундулирующую мембрану, которая отсутствует лишь в самом дистальном конце. 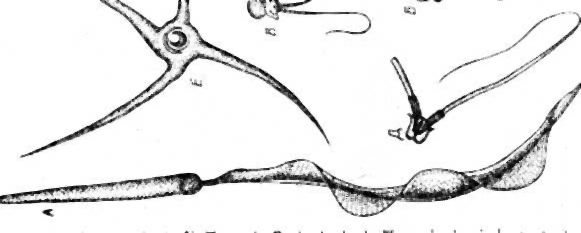  Рис. 4. Сперматозоид тритона (Газарян, Белоусов, 1983). Рис. 4. Сперматозоид тритона (Газарян, Белоусов, 1983).Препарат 4. Сперматозоиды петуха Сперматозоиды петуха являются также очень мелкими жгутиковыми клетками. Передняя часть сперматозоида представлена несколько вытянутой и изогнутой головкой с крупным и компактным ядром. Шейка почти не расширена и переходит в хвостик. При рассмотрении спермы петуха необходимо найти участок, где сперматозоиды лежат поодиночке.  Рис. 5. Сперматозоиды петуха (Новиков, Святенко, 1984). 1 – головка; 2 – хвостик. Занятие 2 Оогенез Оогенезом называется образование женских половых клеток. Яйцеклетка – высокоспециализированная гаплоидная клетка, обладающая потенциальной способностью к развитию. В ее цитоплазме запасаются вещества, необходимые для роста и развития зародыша: структурные белки, ферменты, информационный материал, витамины, соли и др. В отличие от сперматозоида, яйцеклетки увеличивают объем цитоплазмы в десятки, а иногда и в тысячи раз. Основным питательным веществом, накапливаемым в цитоплазме, является желток. По количеству желтка яйцеклетки подразделяются на маложелтковые (олиголецитальные), со средним количеством желтка (мезолецитальные) и многожелтковые (полилецитальные). Иногда яйцеклетки почти лишены желтка (алецитальные). Распределение желтка в яйцеклетке также может быть различным. При равномерном распределении желтка, яйцеклетки называется изо- или гомолецитальными; если он смещен к одному из полюсов –телолецитальными. В случае если желток распределен в яйцеклетке неравномерно, то она приобретает полярность. Желток в большей степени откладывается на одном из полюсов (вегетативном), а на противоположном ему (анимальном) находится цитоплазма и сюда, как правило, смещается и ядро клетки. У членистоногих яйцеклетки полилецитальные, но желток расположен в центральной части цитоплазмы, ядро также расположено в центре (центролецитальные). Яйцеклетки, помимо плазматической оболочки, обычно имеют еще ряд оболочек. Так, сама яйцеклетка вырабатывает первичную или желточную оболочку. Вторичная оболочка формируется фолликулярными клетками, окружающими овулировавшее из яичника яйцо. Третичные оболочки, которые вырабатываются стенками яйцеводов, имеют наиболее сложное строение. Размеры и форма яиц варьируют в пределах животного царства. Как и при сперматогенезе, в оогенезе выделяются те же этапы, но отсутствует период формирования. В период размножения диплоидные оогонии совершают видоспецифичное число митотических деления, а затем переходят в стадию роста. Отличительной особенностью оогенеза является то, что размножающиеся ооциты обладают меньшей митотической активностью. У высших позвоночных пролиферация ооцитов осуществляется только в эмбриогенезе. В период роста в клетке осуществляются одновременно два процесса: в ядре – профаза первого мейотического деления, а в цитоплазме – накопление питательного и информационного материала. Рост ооцита делится на два периода:
Накопление питательного материала в превителлогенезе происходит за счет собственного метаболизма клетки, а в вителлогенезе – за счет поступления его извне. В цитоплазме ооцитов желток запасается в виде гранул и представляет собой смесь компонентов (белки, жиры, углеводы), основным из которых является белок вителлогенин. У большинства позвоночных и некоторых беспозвоночных вителлогенин синтезируется в печени. В период вителлогенеза ооцит увеличивается в размерах в десятки, а иногда и в тысячи раз. Заканчивается оогенез делениями созревания. В результате первого деления происходит редукция числа хромосом, в результате второго – в дочерние клетки расходятся хроматиды. В результате формируются 4 гаплоидных клетки, но только одна из них становится зрелой яйцеклеткой. Итак, оогенез отличается от сперматогенеза рядом признаков:
Типы оогенеза Развитие женских половых клеток может осуществляться в любом участке тела (диффузный оогенез), либо в специализированных органах – яичниках (локализованный оогенез). Диффузный оогенез характерен для примитивных многоклеточных (губки), когда развивающиеся ооциты накапливают питательные вещества путем фагоцитоза близлежащих клеток. При локализованном оогенезе транспорт питательных веществ извне осуществляется специализированными клетками (алиментарный тип) или без участия последних (солитарный тип). При солитарном накоплении питательных веществ все необходимые макромолекулы ооцит получает из полости гонады в виде простых низкомолекулярных соединений. Алиментарный тип оогенеза может быть фолликулярным или нутриментарным. При фолликулярном оогенезе растущий ооцит окружен фолликулярными клетками (соматическими по происхождению), которые вместе с ним образуют функциональную структуру – фолликул. Это наиболее распространенный тип оогенеза, происходящий у всех хордовых. При нутриментарном оогенезе ооцит окружен трофоцитами (клетками-кормилками), связанными с ним цитоплазматическими мостиками. Трофоциты – абортивные половые клетки, имеющие общее происхождение с развивающимся ооцитом. Нутриментарный оогенез описан для высших червей и насекомых. Яичник млекопитающих – это плотный орган, строма которого представлена соединительной тканью. На поперечных срезах различимо корковое вещество (cortex), в котором находятся фолликулы всех стадий развития, и мозговое вещество (medulla), где проходят крупные сосуды яичника. Стенка яйцевых фолликулов образована соединительнотканной текой, в которой разветвляются капилляры, питающие фолликул, и фолликулярным эпителием. Впервые половая клетка устанавливает связь с фолликулярными, когда вступает в период малого роста и превращается в ооцит 1 порядка В это время в ее ядре начинается профаза мейоза, но не завершается, а блокируется на стадии диплотены. Так образуются примордиальные или первичные фолликулы (рис. 6).. Они представляют собой резерв половых клеток, которые возобновляют свое развитие при половом созревании. Располагаются они, как правило, группами в самых поверхностных участках корковой зоны яичника и являются генерацией покоящихся фолликулов. Примордиальные фолликулы порциями вступают в рост в течение всей жизни самки. При инициации развития фолликулярные клетки начинают пролиферировать, из плоских превращаются в кубические, а затем и призматические, и начинают покрывать ооцит первого порядка сначала одним слоем, а затем двумя, тремя и др. Фолликулярная оболочка многослойного фолликула называется зернистой (zonagranulosa), снаружи от нее начинает формироваться соединительнотканная оболочка (tecafolliculi). После того как вокруг ооцита первого порядка завершается формирование одного слоя фолликулярных клеток, начинается формирование блестящей оболочки ооцита (zonapellucida). Эту оболочку пронизывают микровыросты ооцита и макровыросты фолликулярных клеток. Первичный фолликул увеличивается в размерах, как за счет собственного роста, так и за счет увеличения количества слоев фолликулярных клеток. |