методичка БРР. Учебнометодическое пособие для студентов очной и очнозаочной формы обучения по специальности 020201 биология
 Скачать 19.97 Mb. Скачать 19.97 Mb.
|

|
Препарат 1. Яичник кошки (гистологический срез) 1 3 2    Рис. 6. Яичник кошки (Мануилова, 1964). 1 – первичный фолликул; 2 – фолликулярный эпителий; 3 – цитоплазма ооцита. Со временем между развивающимся ооцитом и фолликулярными клетками происходит образование полостей, заполненных жидкостью. На этом этапе фолликул получает название вторичного фолликула. Затем рост ооцита прекращается, а фолликул продолжает расти за счет пролиферации клеток зернистой оболочки и образования полостей. При слиянии полостей в общую полость (antrum), образуется третичный фолликул или граафов пузырек (рис. 7). Граафов пузырек состоит из нескольких слоев клеток. В том месте, где расположен ооцит, фолликулярные клетки образуют яйценосный бугорок.  4 6 Рис. 7. Граафов пузырек из яичника кошки (Мануилова, 1964). 1 – яйцо; 2 – яйценосный бугорок; 3 – блестящая оболочка; 4 – тека; 5 – полость пузырька; 6 – фолликулярный эпителий. Ооцит окружен блестящей оболочкой (zonapellucida) и лучистым венцом (coronaradiata). Полость фолликула ограничена зернистой оболочкой (zonagranulosa). Снаружи фолликул покрыт базальной мембраной и текальной оболочкой, состоящей из внутренней части, содержащей сосуды, и наружной, в состав которой входят стромальные и мышечные клетки. Граафов пузырек на завершающих стадиях своего развития быстро увеличивается в размерах, приближается к поверхности яичника, и его верхушка (стигма) выдается над ней. Впоследствии происходит разрыв cтенки в области стигмы и овуляция ооцита. На месте овулировавшего ооцита, оставшиеся фолликулярные клетки претерпевают железистый метаморфоз и превращаются в желтое тело. Препарат 2. Строение яйцеклетки лягушки Размножение лягушек происходит ранней весной после пробуждения от зимней спячки. Перед овуляцией яйцевые клетки располагаются внутри фолликулов в стенках яичника, а яйцеводы набухают и содержат студенистую массу. Проходя по яйцеводам, яйцеклетки обволакиваются студенистым веществом, поэтому при откладке оказываются покрыты тонкой желточной и студенистой оболочками (рис.8). Последняя оболочка служит защитой для яйца и, попадая в воду, сильно набухает. В воде отложенные яйца достигают размеров в 4-7 раз больших, чем сама яйцеклетка. Такая мощная защитная оболочка выполняет несколько функций: во-первых, является несъедобной для хищников; во-вторых, удерживает яйца около поверхности воды; в-третьих, предохраняет развивающееся яйцо от потери тепла. Неоплодотворенное яйцо лягушки окрашено равномерно в темно-серый цвет, но после оплодотворения черный пигмент скапливается на анимальном полюсе для поглощения солнечных лучей. На всех этапах стадии роста яйцеклетки окружены слоем плоских фолликулярных клеток с ядрами округлой, овальной или палочковидной формы. 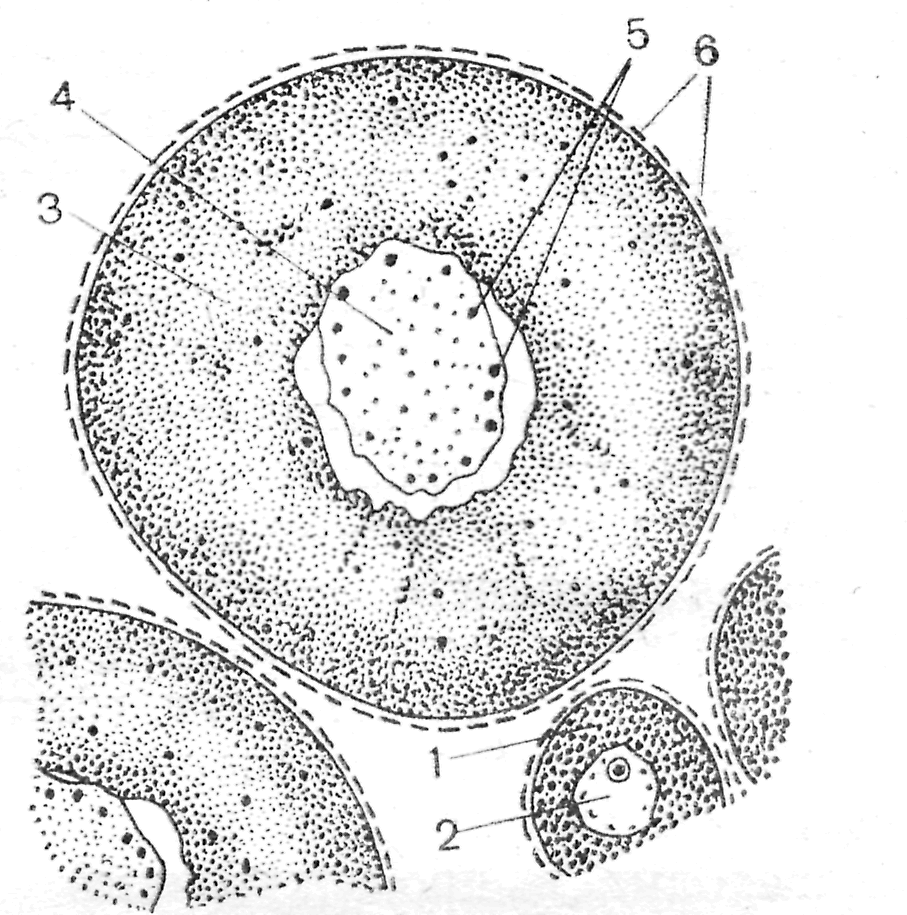 Рис. 8. Яйцеклетка лягушки (Мануилова, 1964). 1 – растущая яйцеклетка; 2 – ядро; 3 – ооцит в фазе большого роста; 4 – ядро; 5 – ядрышки; 6 – ядра фолликулярных клеток. Препарат 3. Строение яйцеклетки беззубки Яйца пластинчатожаберных моллюсков содержат мало желтка, мелки и полярность их выражена слабо. Обычно они имеют диаметр 50-60 мк. Оогенез протекает по солитарному типу, т. е. никаких питающих клеток возле ооцита нет. Ооциты образуются из зачаткового эпителия стенки гонады. По мере роста они начинают выдаваться из стенки и соединяются с нею лишь тонкой ножкой. На свободной поверхности ооцита образуется желточная оболочка. Затем ооцит отделяется от стенки гонады, а на месте его бывшего прикрепления в желточной оболочке остается отверстие, играющее роль микропиле. Помимо желточной оболочки имеется еще вторичная – студенистая оболочка (рис. 9). Препарат следует рассматривать при большом увеличении микроскопа. 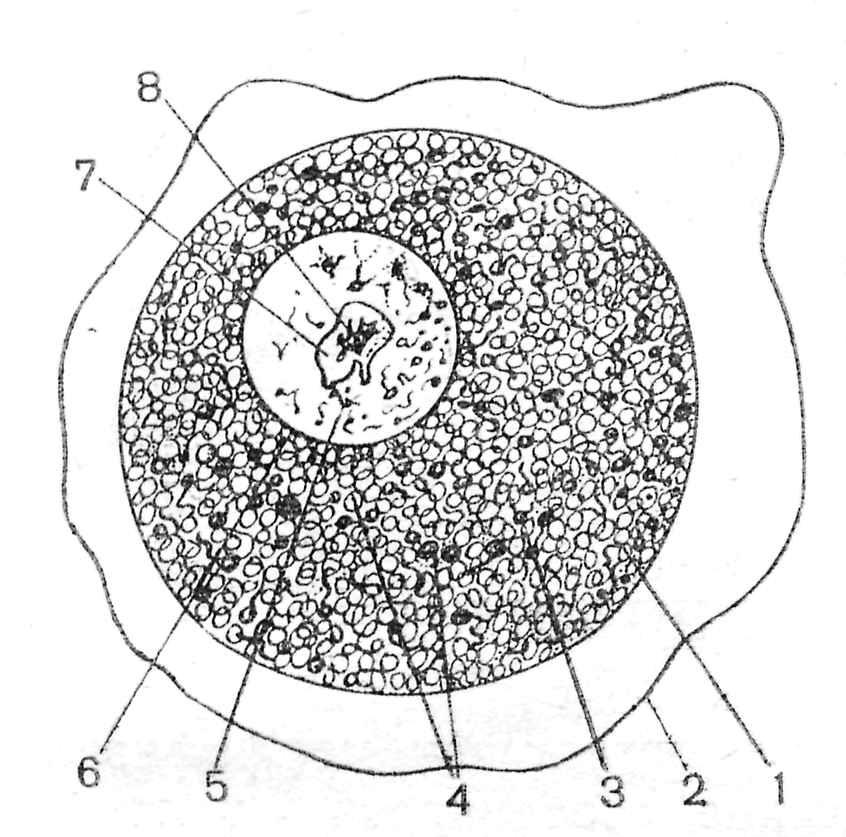 Рис. 9. Яйцеклетка беззубки (Мануилова, 1964). 1 – первичная оболочка; 2 – вторичная оболочка; 3 – цитоплазма; 4 – зерна желтка; 5 – ядро; 6 – ядерная оболочка; 7, 8 – части ядрышка. . Занятие 3 Оплодотворение В зрелых половых клетках практически все жизненные процессы подавлены. Клетки фактически находятся в состоянии анабиоза, и только оплодотворение у большинства видов животных является толчком к дальнейшему развитию. Оплодотворение представляет собой процесс слияния генетического материала двух половых клеток (яйцеклетки и сперматозоида), в результате чего возникает новая особь с потенциями обоих родителей. Таким образом, оплодотворение осуществляет две основные функции: – половую, когда комбинируются гены двух родителей; – репродуктивную, когда создается новый организм. Всякому оплодотворению предшествует процесс осеменения. Осеменение может быть наружным и внутренним. Наружное осеменение свойственно большинству водных животных или тем, кто размножается в воде. У животных, размножающихся на суше, как правило, оплодотворение внутреннее. При осеменении у животных и человека происходит выделение большого количества сперматозоидов. Среда женских половых путей не очень благоприятна для сперматозоидов, и миллионы их погибают, не достигнув яйцеклетки. Дистантное взаимодействие гамет обеспечивает вероятность встречи сперматозоида с яйцеклеткой. При наружном оплодотворении – это видоспецифическое привлечение спермиев своего вида и их активация посредством хемотаксисов. При внутреннем оплодотворении, происходящем (например, у человека), в верхней трети яйцеводов, основная роль в дистантном взаимодействии гамет отводится явлению реотаксиса – способности спермиев двигаться против встречного течения жидкости в маточных трубах. Контактное взаимодействие гамет складывается из нескольких реакций. Первая из них – акросомная. Сначала происходит слияние наружной мембраны акросомы с плазматической мембраной сперматозоида, а затем – разрыв этих мембран, сопровождающийся экзоцитозом содержимого акросомного пузырька. Сперматолизины, являющиеся основным компонентом акросомы, растворяют третичную оболочку яйцеклетки. Благодаря выходу ионов водорода из головки спермия происходит образование акросомного выроста. Акросомный вырост вступает в контакт с наружной поверхностью яйцеклетки – желточной оболочкой. Белок биндин отвечает за специфическую реакцию взаимодействия мембран яйцеклетки и спермия и встроен в головку спермия. Далее происходит активация метаболизма яйцеклетки. В месте проникновения спермия на поверхности яйцеклетки образуется бугорок оплодотворения. Формирование его сопровождается активацией белка актина яйцеклетки. Мембраны акросомной нити и яйцеклетки сливаются, и через образовавшийся цитоплазматический мостик происходит слияние цитоплазмы спермия и ооцита (плазмогамия), а также проникновение сперматического ядра в ооплазму. Участок мембраны сперматозоида встраивается в мембрану яйцеклетки. После контакта сперматозоида с плазматической мембраной яйцеклетки в первые секунды происходит блок полиспермии (у большинства групп животных). В яйцеклетке происходит высвобождение ионов кальция из клеточных депо, а это в свою очередь приводит к кортикальной реакции и образованию оболочки оплодотворения. Полное завершение мейоза без участия сперматозоида, происходит лишь у немногих видов животных (некоторые кишечнополостные, некоторые иглокожие). У большинства видов мейоз не завершается и блокируется в конце профазы (как правило, на стадии диплотены-диакинеза). Это – первый блок мейоза, который у позвоночных может продолжаться годами и снимается во время овуляции. Однако и после этого часто мейоз не доходит до конца – наступает второй блок мейоза, краткосрочный. Для завершения мейоза необходимо снять этот блок активирующим агентом. В роли такого агента выступает сперматозоид. Всех животных можно поделить на группы в зависимости от того, как эти остановки мейоза накладываются на процессы овуляции и оплодотворения:
У большинства животных сперматозоид входит в яйцеклетку целиком, включая и хвостик. Но даже если жгутик и окружающие его митохондрии проникли в яйцеклетку, они не играют никакой роли при дальнейшем развитии. Попав в яйцеклетку, сперматическое ядро поворачивается на 180°, и дистальная центриоль встает впереди него, осуществляя передвижение ядра в ооплазме. Затем сперматическое ядро преобразуется в мужской пронуклеус. В это время в яйцеклетке завершаются деления созревания и пронуклеусы совершают сложные движения (танец пронуклеусов) навстречу друг другу. Оплодотворение яйцеклеток и созревательные деления (на примере аскариды) Препарат 1. Оплодотворение яйцеклетки На рис. 10 представлена схема, иллюстрирующая проникновение сперматозоида в яйцеклетку. Цитоплазма яйцеклетки сильно вакуолизирована. В центре находится ядро с 4 хромосомами (диплоидный набор). В сперматозоиде уже гаплоидный набор с 2 хромосомами. Все последующие препараты необходимо рассматривать при большом увеличении микроскопа.  Рис. 10. Оплодотворение яйцеклетки (Новиков, Святенко, 1984). 1 – сперматозоид; 2 – «конус», или «воспринимающий» бугорок оплодотворения; 5 –ядро яйцеклетки; 6 – пенистая цитоплазма. Препарат 2. Матка аскариды (поперечный срез): выделение первого редукционного тельца (рис. 11, А, Б) После внедрения сперматозоида в яйцеклетку в ней начинаются деления созревания (мейоз), сопровождающиеся выделением редукционных телец. Снаружи матка покрыта покровными фолликулярными клетками, а полость ее заполнена оплодотворенными ооцитами. Цитоплазма ооцита сильно вакуолизирована. Женское ядро уходит к анимальному полюсу клетки и формирует там веретено первого деления созревания. После прохождения конъюгации и репликации ДНК в ядре находятся 2 хромосомы, состоящие из 4 хроматид (тетрады). Оболочка оплодотворения еще не отошла от мембраны ооцита. Сперматическое ядро набухает, в нем еще виден хроматин. Препарат 3. Матка аскариды (поперечный срез): выделение второго редукционного тельца и образование оболочки оплодотворения (рис. 11, В, Г) На препарате видно, что образовалось перивителлиновое пространство, и оболочка оплодотворения отошла от зиготы. Зигота имеет мелкозернистую цитоплазму, в которой лежит деконденсированная головка спермия. На этом этапе деконденсация почти завершилась: темная головка сперматозоида почти не видна, а область хроматина мужского ядра большего диаметра и более плотная, чем на предыдущем препарате. На периферии зиготы видно веретено второго деления созревания. Каждая хромосома состоит уже из 2 хроматид (диада). На мембране ооцита располагается первое редукционное тельце. Кроме того, на препарате различима тонкая, но темная оболочка, в виде ободка, покрывающего оболочку оплодотворения. Она называется кутикулой. Это третичная оболочка, она формируется после оплодотворения и секретируется клетками матки. Позднее эта оболочка становится более толстой и прочной. Ее функция – защита эмбриона аскариды от воздействия неблагоприятных факторов внешней среды. Препарат 4. Матка аскариды (поперечный срез): стадия пронуклеусов и первое деление дробления (рис. 12, А) Последующие этапы процесса оплодотворения (стадию пронуклеусов, веретено деления зиготы, стадию двух бластомеров) можно найти на одном препарате. Стадия двух пронуклеусов. После завершения второго редукционного деления гаплоидные родительские геномы покрываются ядерными оболочками – формируются мужской и женский пронуклеусы, которые начинают сближаться, двигаясь по цитоплазме зиготы по сложной траектории. На большом увеличении в центре клетки видны два пронуклеуса. Часто внутри пронуклеусов виден ядерный материал. Он находится в спирализованном состоянии. Перивителлиновое пространство увеличилось в размерах значительно, на яйцеклетке и на внутренней стороне оболочки оплодотворения просматриваются два редукционных тельца. Оболочка оплодотворения покрыта толстой кутикулой. Веретено деления зиготы (рис. 12, А). Сперматозоид вносит в яйцеклетку не только свое ядро, но и центриоль, которая делится и формирует веретено деления зиготы. В центре клетки часто видна метафазная пластинка веретена деления зиготы и даже само веретено. Оно образовано микротрубочками, отходящими от центриолей. Каждая хромосома состоит из двух хроматид – всего 4 хроматиды. Препарат 5. Матка аскариды (поперечный срез): стадия двух и четырех бластомеров. Стадия образования двух бластомеров (рис. 12, Б, В, Г). На поперечном срезе матки аскариды часто заметны эмбрионы на стадии двух бластомеров. Стадия четырех бластомеров (рис. 12, Д, Е, Ж) При втором делении один бластомер делится меридиональной, а второй – горизонтальной бороздой. Расположение бластомеров на этой стадии описывается как «Т- образная фигура». Через нее можно провести только одну плоскость симметрии, поэтому дробление аскариды на стадии 4 бластомеров описывается как билатеральное. 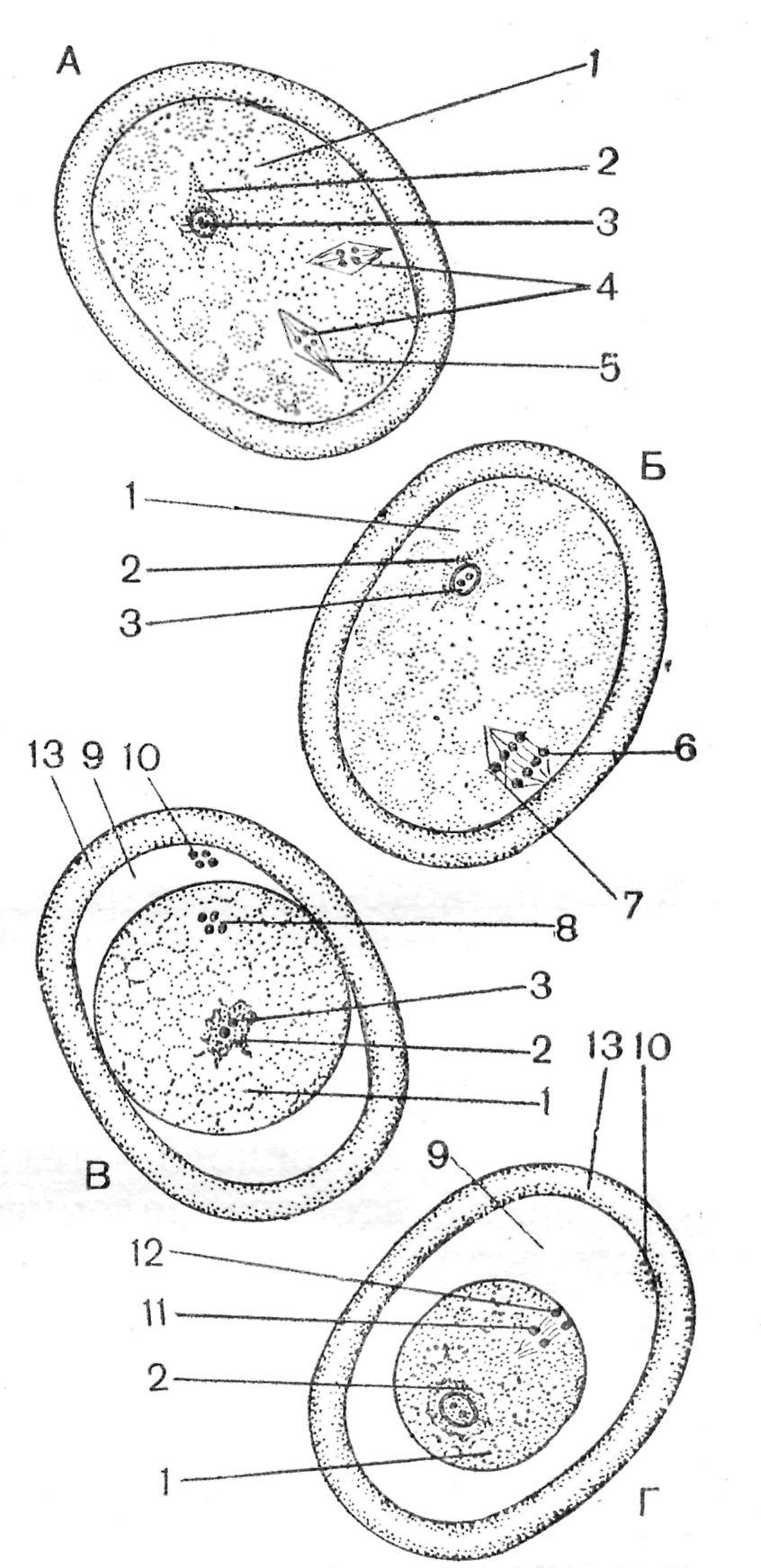 Рис. 11. Схема созревательных делений в яйцеклетках аскариды (Новиков, Святенко, 1984). 1 – пенистая цитоплазма; 2 – сперматозоид; 3 – хромосомы; 4 – хромосомная структура женского ядра в виде двух тетрад; 5 – центральное ахроматиновое веретено; 6, 7 – хромосомы; 8 – хромосомная структура женского ядра в виде диады; 9 – перивителлиновое пространство; 10 – первое редукционное тельце; 11, 12 –расходящиеся хроматиды; 13 – оболочка яйцеклетки. 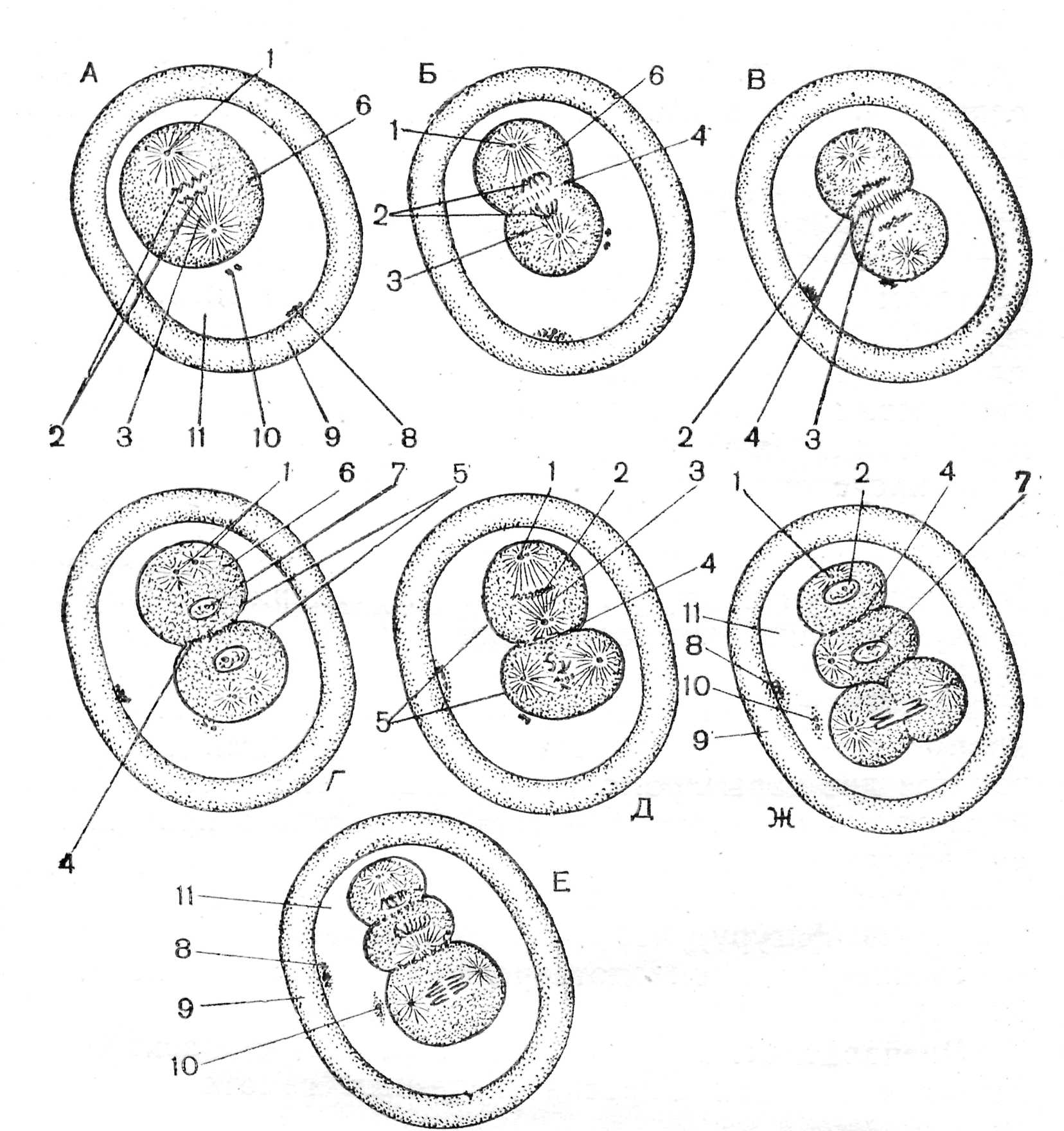 Рис. 12. Веретено деления зиготы и начальные этапы дробления (Новиков, Святенко, 1984). А – веретено деления зиготы; Б, В, Г – последовательные этапы образования двух бластомеров; Д, Е – последовательные этапы образования четырех бластомеров; Ж – «Т- образная фигура». 1 – центриоль; 2 – хромосомы; 3 – веретено деления; 4 – начальная перетяжка зиготы; 5 – 2 бластомера; 6 – цитоплазма бластомеров; 7 – ядро бластомера; 8 – первое редукционное тельце; 9 – оболочка; 10 – второе редукционное тельце; 11 – перивителлиновое пространство. Занятие 4 Дробление и бластуляция Дробление – самый ранний период развития зародыша, во время которого оплодотворенное яйцо делится на большое количество клеток. В период дробления митотический цикл имеет малую продолжительность, в нем не наблюдаются пре- и постсинтетические фазы, отсутствует постмитотический рост клеток. Умножение числа мелких клеток приводит к огромному увеличению их общей площади. В ходе митотического дробления в геометрической прогрессии увеличивается содержание и синтез ДНК, восстанавливается ядерно-цитоплазматическое соотношение, которое нарушается в период вителлогенеза ооцита. Дочерние клетки, образующиеся в период дробления, называются бластомерами, а делящие их плоскости – бороздами дробления. Борозды, проходящие через анимальный и вегетативный полюса зиготы, называются меридиональными. Борозды, проходящие перпендикулярно к полярной оси зиготы, проходящей через ее середину, называются экваториальными. Борозды, смещенные к анимальному полюсу, получили название широтных; а борозды, проходящие параллельно поверхности зародыша, называются тангенциальными. На начальных этапах дробления происходит синхронное деление всех бластомеров. Впоследствии появляется и нарастает асинхронность. На расположение ядер, веретен дробления и бластомеров влияют количество и расположение желтка в цитоплазме яйцеклеток. Желток, будучи необходимым питательным материалом для развития зародыша, является и пассивным балластом, задерживающим деление зиготы и бластомеров. Чем больше желточных гранул, тем медленнее делится эта часть цитоплазмы. Правила зависимости между расположением желтка и положением ядер и веретен были сформулированы немецким ученым О. Гертвигом на основе правил, предложенных ранее ботаником Ю. Саксом для растительных меристем. Суть этих правил состоит в следующем:
На таблице 1 показана связь типов дробления яиц с количеством и распределением в них желтка. Каждый тип дробления следует рассматривать на конкретном примере: полное, более или менее равномерное встречается у морского ежа; полное неравномерное – у амфибий и хрящевых ганоидов; неполное дискоидальное – у костистых рыб, рептилий и птиц. Яйцеклетки с небольшим и средним количеством желтка (олиго- и мезолецитальные) претерпевают полное (голобластическое) дробление. При этом содержимое зиготы входит в состав бластомеров и зародыш развивается за счет внутриклеточной утилизации желтка. Полное дробление может быть равномерным (олиголецитальные яйца) и неравномерным (мезолецитальные яйца). Яйцеклетки с большим количеством желтка (полилецитальные) претерпевают неполное (меробластическое) дробление. Меробластический тип дробления характеризуется тем, что зародыш развивается лишь из небольшой части яйцеклетки, а остальная часть ее – питательный материал. Неполное дробление может быть дискоидальным (телолецитальные яйцеклетки с большим количеством желтка) и поверхностным (центролецитальные яйцеклетки). Характер дробления зависит не только от количества и расположения желтка, но и от ряда других факторов, в частности, от расположения кортикальных гранул в цитоплазме яйцеклетки. Так, дробление может быть радиальным, спиральным, билатеральным, анархичным и т.п. При радиальном дроблении плоскости последовательных дроблений проходят перпендикулярно друг к другу, и бластомеры располагаются симметрично относительно любой плоскости, проходящей через анимально-вегетативную ось яйца. При спиральном дроблении происходит смещение (поворот) бластомеров относительно оси яйца. Это вызывает скручивание веретена деления в анафазе. Плоскости веретен образуют спирали или участки спиралей, и бластомеры располагаются не правильными рядами, а более или менее чередуются друг с другом. Заключительный этап дробления – бластуляция характеризуется появлением ряда новых морфологических особенностей зародыша – бластодермы, первичной полости тела (бластоцели); новых биохимических функций в зародышевых клетках: начало работы зародышевого генома, начало синтеза РНК, белка. Бластоцель – это первый отсек внутренней среды организма, отличается по своему ионному составу от окружающей среды. Изоляция бластоцели от наружной среды обеспечивается формированием на боковых поверхностях соседних бластомеров специализированных плотных межклеточных контактов, непроницаемых для ионов. Проникновение ионов в бластоцель и обратно осуществляется через клетки бластодермы. |