физиология ДЫХАНИЕ. Учебнометодическое пособие физиология дыхания
 Скачать 11.5 Mb. Скачать 11.5 Mb.
|

Диффузия газов через аэрогематический барьерВ организме газообмен кислорода и углекислого газа, а так же других газообразных продуктов происходит с помощью диффузии. Диффузия газов через альвеолокапиллярную мембрану легких осуществляется в два этапа. На первом этапе диффузионный перенос газов происходит по концентрационному градиенту через тонкий аэрогематический барьер (его толщина равна около 1мкм). На втором этапе происходит связывание газов в крови легочных капилляров. Диффузия газов осуществляется в соответствии с градиентом парциальных давлений газов и описывается законом Фика: Q газа = S DK (P1-P2) /T Где Q газа - объем газа, проходящий через ткань в единицу времени, S- площадь ткани, DK- диффузионный коэффициент газа, P1-P2 - градиент парциального давления газа, Т - толщина барьера ткани.  Рисунок 8. Строение аэрогематического барьера 1-сурфактант, 2-эпителий альвеол, 3-интерстициальное пространство, 4-эндотелий капилляров ,5-плазма крови, 6-эритроцит Как видно из приведенной формулы. Диффузия газа зависит от градиента давлений этого газа по обе стороны барьера, следовательно, нас интересуют парциальные давления кислорода и углекислого газа в альвеолярном воздухе и напряжения этих газов в венозной крови. Все эти цифры представлены в таблице 2. Отметим лишь, что в альвеолярном воздухе часть общего давления ( 47 мм рт.ст.) приходится на пары воды, значит давление «сухого» воздуха = 760 – 47 = 713 мм рт.ст. Альвеолярный воздух обогащен углекислым газом, кислорода в нем не 21, а 14%, следовательно парциальное давление кислорода в нем составит 14 % от 713 = 100 мм рт.ст. В венозной крови легочных капилляров напряжение кислорода = 40 мм рт.ст. Градиент давлений, обеспечивающий диффузию кислорода равен 100 – 40 = 60 мм рт.ст. Что касается диффузии СО2 из венозной крови в альвеолы, то даже сравнительно небольшого градиента РСО2 (6-10мм.рт.ст.) для этого оказывается вполне достаточно, поскольку растворимость углекислого газа в 20-25 раз больше, чем кислорода. Именно поэтому после прохождения венозной крови через легочные капилляры РСО2 в ней оказывается почти равным альвеолярному (около 40 мм.рт. ст.). Для кислорода Р1- Р2 = 60 мм рт.ст Для углекислого газа Р1- Р2 = 6 мм рт.ст Ещё раз необходимо подчеркнуть, что постоянная скорость диффузии, как кислорода, так и углекислого газа через аэрогематический барьер определяются достаточно стабильным составом альвеолярного газа во время вдоха и выдоха.Капилляры легкихФункции газообмена в легких и насыщение крови кислородом осуществляется с участием сосудов малого круга кровообращения. Стенки ветвей легочной артерии тоньше, чем стенки такого же калибра артерий большого круга кровообращения. Сосудистая система легких очень податлива и способна легко растягиваться. В систему легочной артерии поступает сравнительно большой объем крови (6 литров/мин) из правого желудочка, а давление в малом круге низкое - 15-20 мм рт. ст., потому, что сосудистое сопротивление примерно в 10 раз меньше, чем в сосудах большого круга кровообращения. Сеть альвеолярных капилляров не сравнима с организацией капиллярного русла других органов. Отличительными чертами капиллярного русла легких являются 1) малая величина капиллярных сегментов, 2) их обильная взаимосвязь, что формирует петлистую сеть, 3) высокая плотность отдельных капиллярных сегментов на единицу площади альвеолярной поверхности, 4) низкая скорость кровотока. Капиллярная сеть в стенках альвеол настолько плотная, что некоторые физиологи рассматривают ее как сплошной слой движущейся крови. Площадь поверхности капиллярной сети близка площади поверхности альвеол (80 м2), в ней содержится около 200 мл крови. Диаметр альвеолярных кровеносных капилляров колеблется в пределах 8.3 - 9.9 мкм, а диаметр эритроцитов - 7.4 мкм. Таким образом, эритроциты плотно прилегают к стенкам капилляров. Эти особенности кровоснабжения легких создают условия для быстрого и эффективного газообмена, в результате которого происходит уравновешивание газового состава альвеолярного воздуха и артериальной крови. Взгляните еще раз на таблицу 2 и отметьте, что напряжение кислорода в артериальной крови становится равным 100, а углекислого газа – 40 мм рт. ст. Транспорт кислорода кровьюБольшая часть кислорода в организме млекопитающих переносится кровью в виде химического соединения с гемоглобином. Свободно растворенного кислорода в крови всего 0.3%. Реакцию оксигенации, превращение дезоксигемоглобина в оксигемоглобин, протекающую в эритроцитах капилляров легких можно записать следующим образом: Нв + 4О2 ⇄ Нв(О2)4 Эта реакция протекает очень быстро – время полунасыщения гемоглобина кислородом около 3 миллисекунд. Гемоглобин обладает двумя удивительными свойствами, которые позволяют ему быть идеальным переносчиком кислорода. Первое – это способность присоединять кислород, а второе – отдавать его. Оказывается способность гемоглобина присоединять и отдавать кислород зависит от напряжения кислорода в крови. Попробуем изобразить графически зависимость количества оксигенированного гемоглобина от напряжения кислорода в крови, и тогда нам удастся выяснить: в каких случаях гемоглобин присоединяет кислород, а в каких отдает. Гемоглобин и оксигемоглобин неодинаково поглощают световые лучи, поэтому их концентрацию можно определить спектрометрическими методами. График, отражающий способность гемоглобина присоединять и отдавать кислород называется «Кривая диссоциации оксигемоглобина». По оси абсцисс на этом графике отложено количество оксигемоглобина в процентах ко всему гемоглобину крови, по оси ординат – напряжение кислорода в крови в мм рт. ст.  Рисунок 9А. Кривая диссоциации оксигемоглобина в норме Рассмотрим график в соответствии с этапами транспорта кислорода: самая высокая точка соответствует тому напряжению кислорода, которое наблюдается в крови легочных капилляров – 100 мм рт.ст. (столько же, сколько и в альвеолярном воздухе). Из графика видно, что при таком напряжении весь гемоглобин переходит в форму оксигемоглобина – насыщается кислородом полностью. Попробуем рассчитать, сколько кислорода связывает гемоглобин. Один моль гемоглобина может связать 4 моля О2 , а 1грамм Нв связывает 1,39 мл О2 в идеале, а на практике 1,34 мл . При концентрации гемоглобина в крови, например, 140 г/литр количество связанного кислорода составит 140 × 1,34 = 189,6 мл/литр крови. Количество кислорода, которое может связать гемоглобин при условии его полного насыщения, называется кислородной емкостью крови (КЕК). В нашем случае КЕК = 189,6 мл. Обратим внимание на важную особенность гемоглобина – при снижении напряжения кислорода в крови до 60 мм рт.ст, насыщение практически не изменяется – почти весь гемоглобин присутствует в виде оксигемоглобина. Эта особенность позволяет связывать максимально возможное количество кислорода при снижении его содержания в окружающей среде (например, на высоте до 3000 метров). Кривая диссоциации имеет s – образный характер, что связано с особенностями взаимодействия кислорода с гемоглобином. Молекула гемоглобина связывает поэтапно 4 молекулы кислорода. Связывание первой молекулы резко увеличивает связывающую способность, так же действуют и вторая, и третья молекулы. Этот эффект называется кооперативное действие кислорода Артериальная кровь поступает в большой круг кровообращения и доставляется к тканям. Напряжение кислорода в тканях, как видно из таблицы 2, колеблется от 0 до 20 мм рт. ст., незначительное количество физически растворенного кислорода диффундирует в ткани, его напряжение в крови снижается. Снижение напряжения кислорода сопровождается диссоциацией оксигемоглобина и освобождением кислорода. Освободившийся из соединения кислород переходит в физически растворенную форму и может диффундировать в ткани по градиенту напряжения.. На венозном конце капилляра напряжение кислорода равно 40 мм.рт.ст, что соответствует примерно 73% насыщения гемоглобина. Крутая часть кривой диссоциации соответствует напряжению кислорода обычному для тканей организма – 35 мм рт.ст и ниже. Таким образом, кривая диссоциации гемоглобина отражает способность гемоглобина присоединять кислород, если напряжение кислорода в крови высоко, и отдавать его при снижении напряжения кислорода. Переход кислорода в ткани осуществляется путем диффузии, и описывается законом Фика, следовательно зависит от градиента напряжений кислорода. Можно узнать, сколько кислорода извлекается тканью. Для этого нужно определить количество кислорода в артериальной крови и в венозной крови, оттекающей от определенной области. В артериальной крови, как нам удалось вычислить (КЕК) содержится 180-200 мл. кислорода. Венозная кровь в состоянии покоя содержит около 120 мл. кислорода. Попробуем рассчитать коэффициент утилизации кислорода: 180 мл. 120 мл. = 60 мл.- это количество извлеченного тканями кислорода, 60мл./180 100 = 33%. Следовательно, коэффициент утилизации кислорода равен 33% (в норме от 25 до 40%). Как видно из этих данных, не весь кислород утилизируется тканями. В норме в течение одной минуты к тканям доставляется около 1000 мл. кислорода. Если учесть коэффициент утилизации, становится ясно, что ткани извлекают от 250 до 400 мл. кислорода в минуту, остальной кислород возвращается к сердцу в составе венозной крови. При тяжелой мышечной работе коэффициент утилизации повышается до 50 – 60 %. Однако количество кислорода, которое получают ткани, зависит не только от коэффициента утилизации. При изменении условий во внутренней среде и тех тканях, где осуществляется диффузия кислорода, свойства гемоглобина могут измениться. Изменение свойств гемоглобина отражается на графике и называется «сдвиг кривой». Отметим важную точку на кривой – точка полунасыщения гемоглобина кислородом наблюдается при напряжении кислорода 27 мм рт. ст., при таком напряжении 50 % гемоглобина находится в форме оксигемоглобина, 50% в виде дезоксигемоглобина, следовательно 50 % связанного кислорода – свободно (примерно 100мл/л). Если в ткани увеличивается концентрация углекислого газа, ионов водорода, температура, то кривая сдвигается вправо. В этом случае точка полунасыщения переместится к более высоким значениям напряжения кислорода - уже при напряжении 40 мм рт. ст. будет освобождено 50 % кислорода (рисунок 9Б). Интенсивно работающей ткани гемоглобин отдаст кислород легче. Изменение свойств гемоглобина обусловлены следующими причинами: закисление среды в результате увеличения концентрации углекислого газа действует двумя путями 1) увеличение концентрации ионов водорода способствует отдаче кислорода оксигемоглобином потому, что ионы водорода легче связываются с дезоксигемоглобином, 2) прямое связывание углекислого газа с белковой частью молекулы гемоглобина уменьшает ее сродство к кислороду; увеличение концентрации 2,3-дифосфоглицерата, который появляется в процессе анаэробного гликолиза и тоже встраивается в белковую часть молекулы гемоглобина и снижает его сродство к кислороду. Сдвиг кривой влево наблюдается, например, у плода, когда в крови определяется большое количество фетального гемоглобина. 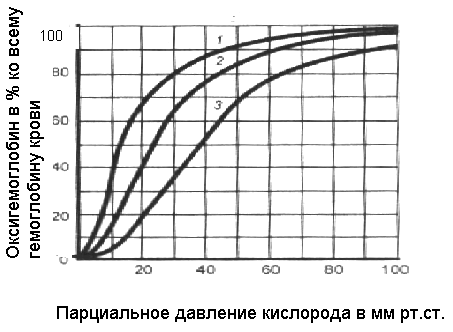 Рисунок 9 Б. Влияние изменения параметров внутренней среды на кривую диссоциации оксигемоглобина 1- при низком содержании СО2 , увеличении рН, снижении температуры, снижении концентрации 2-3 ДФГ 2- норма, 3- при высоком содержании СО2 , низком рН, высокой температуре, высокой концентрации 2-3 ДФГ Транспорт углекислого газаПеренос СО2 из клеток тканей в кровь тоже происходит путем диффузии, т.е. в силу разности напряжений СО2 по обе стороны гемато-паренхиматозного барьера, поскольку среднее значение СО2 в артериальной крови составляет около 40 мм.рт.ст., в то время как в клетках эта величина достигает 60 мм.рт.ст. Локально парциальное давление углекислого газа, а следовательно и скорость его диффузии, определяются в значительной мере скоростью продукции СО2 в клетках (т.е. интенсивностью окислительных процессов в том или ином органе).  |