Учебное пособие для ординаторов по специальности Функциональная диагностика
 Скачать 2.86 Mb. Скачать 2.86 Mb.
|

|
ТЕХНИКА И МЕТОДИКА ЭНМГ. ЭНМГ ОБОРУДОВАНИЕ. Электронейромиограф – прибор для активации, регистрации и изучения биопотенциалов мышц и нервов. Используемые в последнее время компьютерные электронейромиографы имеют ряд преимуществ в регистрации, обработке, накоплении сигналов. Современные электронейромиографы состоят из электродов, соединительных проводов, предусилителъного блока с аналого-цифровым преобразователем, усилителей, дисплея, компьютерного блока и стимулятора. Электроды В более общем виде электроды делятся нарегистрирующие, стимулирующие и заземляющие. Регистрирующие электроды предназначены для регистрации биопотенциалов. Они могут быть пластинчатыми (поверхностными, или накожными), игольчатыми и проволочными. Пластинчатые электроды, как правило, используют для отведения биоэлектрической активности с поверхности кожи, поэтому их часто называют поверхностными, или накожными электродами. Поверхностные электроды позволяют отводить биопотенциалы с поверхности кожи над двигательной точкой мышцы и пи над нервом. Форма пластинчатых электродов может быть круглой или прямоугольной, площадь 30-100 кв. мм, толщина от 1 до 3 мм. Для взрослых используются электроды размером 5x10 мм с фиксированным расстоянием 20 мм между их центрами. Электроды изготавливаются из нержавеющей стали или, лучше, из серебра (хлорсеребряные) для уменьшения их поляризации, т.е. накопления на их поверхности электрического заряда, и минимизации помех при регистрации ЭНМГ. Поверхностные электроды используются при регистрации поверхностной и стимуляционной ЭМГ. В стимуляционной ЭМГ используют как биполярные, так и монополярные регистрирующие электроды. Монополярные электроды предназначены для монополярной регистрации, когда активный электрод находится над двигательной точкой мышцы или над нервом, т.е. в активной зоне генерации потенциала, а референтный электрод отнесен от точки регистрации на некоторое расстояние (на сухожилие мышцы или на 2-3 см от точки проекции нерва), т.е. находится вне зоны генерации потенциала. Согласно договоренности, активным электродом считается тот электрод, под которым разность потенциалов является отрицательной. Условным обозначением активного электрода является знак «-», а референтного - знак «+». Обе пластины накожного биполярного регистрирующего электрода для регистрации потенциала сенсорного нерва обычно располагают над нервом с межэлектродным расстоянием 4 см. Игольчатые регистрирующие электроды предназначены для отведения биопотенциалов в непосредственной близости от источника генерации потенциала - от мышечных волокон, двигательных единиц мышцы, нерва. Игольчатые электроды могут быть моно- и биполярные. Стимулирующие электроды, как и регистрирующие, могут быть пластинчатыми, игольчатыми и, кроме того, кольцевидными. Пластинчатые электроды универсальны для регистрации и стимуляции. Для стимуляции достаточно глубоко расположенных нервных стволов используют вилочковые электроды. Для стимуляции обычно используют монополярные игольчатые электроды. Кольцевидные электроды используют для стимуляции чувствительных нервов пальцев. Заземляющие электроды предназначены для выравнивания потенциала прибора и тела пациента с целью уменьшения синфазного напряжения помехи на входах усилителя. Хороший контакт заземляющего электрода с кожей (малое переходное сопротивление) снижает помехи в записи ЭМГ. Заземляющие электроды могут быть пластинчатыми или ленточными, последние удобны при наложении вокруг конечности. Усилитель Визуализация биопотенциалов мышц и нервов возможна на экране катодно-лучевой трубки. Однако для осциллоскопа необходим сигнал с величиной, измеряемой в вольтах, а амплитуда ЭНМГ сигналов в тысячи и миллион раз меньше. Усилитель электромиографа позволяет низкоамплитудные регистрируемые сигналы, измеряемые в микровольтах, усилить в 1000-1000000 раз. Этот процесс осуществляется в несколько этапов. Первый этап - это предусиление, которое позволяет усилить сигнал в 500 раз. На последующих этапах сигнал усиливается в 2-2000 раз. Имеется возможность регулировать коэффициент усиления с лицевой панели ЭНМГ или с клавиатуры компьютера путем изменения так называемого уровня чувствительности с 0.5 мкВ/деление (мкВ/дел) до 20 мВ/дел. Канал усилителя может находиться в 3 состояниях: включенном (режим измерения - «measure»); выключенном (режим - «Gnd»), когда в схеме усилителя входные контакты закорачиваются (соединяются между собой и заземляющим электродом); тестирующем (режим - «Cal»), когда в схеме усилителя на входы подается тестовый калибровочный сигнал прямоугольной формы величиной 10 или 100 мкВ. Величина калибровочного сигнала проверяется на дисплее по соответствию с выставленной чувствительностью усилителей, например, при чувствительности 5 мкВ/дел калибровочный сигнал в 10 мкВ будет занимать высоту, равную 2 делениям. В усилительном блоке могут располагаться несколько каналов усиления в точном соответствии с числом каналов электромиографа. Количество каналов в современных электромиографах может быть от 1 до 8. Характеристики чувствительности усилителя, полосы пропускания частот можно устанавливать на разных каналах разные или одинаковые - в зависимости от методики исследования. Во входном гнезде каждого канала усилителя имеются 2 входных контакта (первый и второй). Их нумерация жестко закреплена: активный электрод должен подсоединяться к первому контакту, а референтный - ко второму. Это позволяет получать на экране негативную фазу потенциалов всегда с отклонением вверх от изолинии, а позитивную вниз от изолинии. Дисплей Осциллоскоп (дисплей, или визуализатор) является составляющей частью электромиографа и позволяет увидеть регистрируемые сигналы мышц и нервов на экране. В компьютерных электромиографах функцию дисплея выполняет монитор компьютера. Компьютерный блок Компьютерный блок значительно облегчает проведение исследования и анализ зарегистрированных сигналов. Компьютер электромиографа позволяет управлять с одной клавиатуры рядом процессов: выбирать канал усилителя, устанавливать чувствительность, граничные низкие и высокие частоты каналов усилителя, проверять межэлектродное сопротивление, подавать калибровочный сигнал, накапливать и обрабатывать регистрируемые сигналы, управлять стимулятором и печатающим устройством. Оснащение электромиографа компьютером позволило значительно облегчить процесс накопления ЭНМГ потенциалов (acquire) и хранить их в оперативной памяти компьютера (store). Это значительно сокращает время обследования больного, так как после окончания обследования пациента записанные потенциалы можно просмотреть, выбрать необходимые для анализа и, после удаления ненужной излишней информации, обработать. Электромиографические кривые можно запомнить на жесткий диск компьютера, т.е. архивировать, и использовать эту информацию при динамическом наблюдении. При необходимости иметь документальную копию картина сигналов на экране может быть распечатана на принтере. Стимулятор Электростимулятор позволяет раздражать нервы на протяжении и регистрировать так называемые вызванные потенциалы, т.е. те, которые возникают искусственно в исследовательских целях. Работа стимулятора синхронизирована с монитором для того, чтобы вызванные потенциалы мышц и нервов предъявлялись на экране в одном и том же месте. Стимулятор может генерировать как постоянный ток, так и импульсный ток. В ЭНМГ исследовании в основном используют импульсный ток. Стимулятор генерирует импульсы прямоугольной формы. В электростимуляторах ряда фирм имеется два и более изолированных стимуляционных выхода, позволяющих давать одновременно раздражения в нескольких участках нерва на его протяжении с разной интенсивностью и задержкой. Эта техническая возможность необходима при исследовании прямых и рефлекторных ответов методом коллизии и облегчения. Артефакты и их устранение Электронейромиографические артефакты – потенциалы биологического и небиологического (физического) происхождения, не связанные с активностью мышц и нервов. К физическим артефактам относят: сетевую наводку частотой 50 Гц, шум электродов, артефакты генераторов электромагнитных полей, артефакт искусственного водителя ритма, радиопомехи, артефакты движения и др. К биологическим артефактам относят ЭКГ. Часто встречающимся артефактом является сетевая наводка 50 Гц. Она представляет собой синусоидальное искажение изолинии частотой 50 ГЦ (рис. 43). При большой развертке (1-5 мс/дел) синусоидальную форму наводки трудно идентифицировать, поэтому рекомендуется использовать скорость развертки 20-50 мс/дел. Длительность позитивной и негативной фазы сетевой наводки 50 Гц составляет 10 мс. Звукового эффекта сетевая наводка 50 Гц не имеет. Причины появления 50 Гц наводки и возможные меры ее устранения указаны в таблице 11. Шум электродов - артефакт, проявляющийся высокочастотной низкоамплитудной махристой активностью, которая представлена утолщенной зазубренной нулевой линией амплитудой 3-5 мкВ. Для устранения артефакта провести электролитическую обработку электродов или заменить электрод. Артефакты генераторов электромагнитных полей могут проявляться высокочастотными прерывистыми колебаниями достаточной амплитуды и симулировать комплексы повторяющихся разрядов, регистрируемых игольчатым электродом. Артефакт искусственного водителя ритма и артефакт ЭКГ появляется на ЭМГ при исследовании мышц в непосредственной близости от сердца. Артефакт движения электрода представлен резким высокоамплитудным скачком изолинии с искажением ЭНМГ потенциалов. Появляется артефакт при быстром сокращении мышцы или движении. Для уменьшения артефакта при записи ЭМГ движения необходимо выставить фильтры высокой частоты на 50 Гц. Радиопомехи - высокочастотные, меняющиеся по амплитуде сигналы в соответствии с интенсивностью звука, воспроизводимого динамиком электромиографа. Причиной может явиться близко расположенный радиопередающий центр. Устранить радиопомехи возможно экранированием помещения. ПОНЯТИЕ ДВИГАТЕЛЬНОЙ ЕДИНИЦЫ Нервно-мышечная система представляет собой функционально тесно связанный комплекс скелетных мышц и периферических образований нервной системы: мотонейронов и их аксонов. Функциональным элементом системы является двигательная единица (ДЕ).Данное понятие впервые было введено Шеррингтоном Х.С. (1925) и до сих пор является физиологической основой ЭМГ. Под двигательной единицей подразумевают комплекс, состоящий из двигательной клетки, ее аксона и группы иннервируемых этим аксоном мышечных волокон. Все элементы одной ДЕ функционально одинаковы и действуют по принципу "все или ничего", когда каждый импульс, превышающий определенный порог, приводит к сокращению всех мышечных волокон одной ДЕ. Территории, занимаемые ДЕ, и количество мышечных волокон в них зависят от размера мышцы, ее функции. Анатомически в одном мышечном пучке могут находиться мышечные волокна от разных ДЕ. Мышечные волокна разного типа располагаются мозаично по всему поперечнику мышцы, и рядом может лежать не более 2-3 мышечных волокон одного гистохимического типа (DubovitzV., BrookeM.H., 1973; BuchtalF., SchmalbruchH., 1980). Таким образом, в области одного пучка представлены мышечные волокна разных ДЕ (Гаусманова-Петрусевич И., 1971). В то же время ДЕ никогда не образует своих разветвлений в замкнутой территории, а распространяет их на соседние области (Рис. 1). 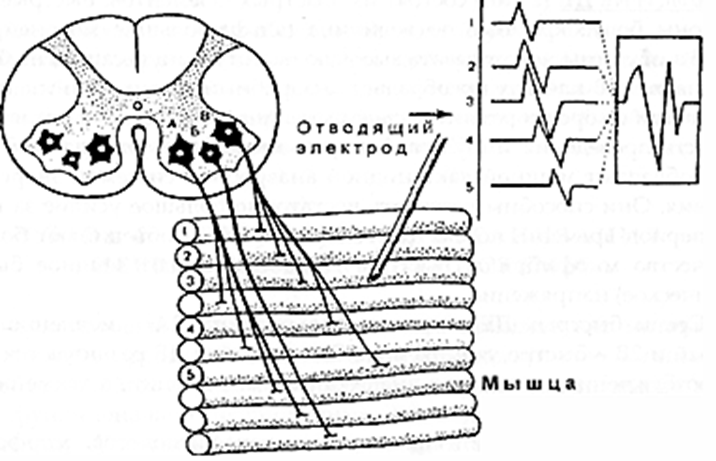 Рис.1 Схема ДЕ. Принцип генерации потенциала двигательной единицы (ПДЕ). А, Б, В - мотонейроны переднего рога спинного мозга; 1-5 - мышечные волокна, относящиеся к территории иннервации мотонейрона В (Л.О. Бадалян, И.А. Скворцов, 1986). Как видно из схемы, зоны отдельных ДЕ перекрываются, но каждая имеет свою специализацию и содержит мышечные волокна одного типа. В функциональном плане ДЕ можно разделить на два основных типа: быстрые и медленные. Существуют и переходные формы. Медленные ДЕ (1 тип) включают медленный мотонейрон, медленный аксон, медленные мышечные волокна. Медленные мотонейроны малые по величине (альфа-малые мотонейроны), имеют высокую возбудимость, низкую частоту генерации импульса, высокую выносливость, не утомляемость. Обмен в них преимущественно аэробный. Аксон этих клеток тонкий, слабо миелинизирован, и скорость проведения возбуждения по нему невелика, но он более возбудим при непрямой электрической стимуляции. Медленные мышечные волокна тоньше, состоят из меньшего количества миофибрилл и поэтому развивают меньшее усилие, однако они более выносливые и могут длительное время давать стабильное напряжение, имеют богатую сеть капилляров, что обеспечивает высокую степень окисления. В целом, медленные ДЕ обеспечивают длительное (тоническое) напряжение мышцы. Быстрые ДЕ (2 тип) состоят из "быстрых" элементов. Быстрые мотонейроны более крупные по величине (альфа-большие мотонейроны), менее возбудимы, могут давать высокую частоту импульсации, но быстро истощаются. В клетках преобладает анаэробный обмен. Они имеют толстый аксон с хорошо развитым слоем миелина, что обеспечивает высокую скорость проведения импульса. Быстрые мышечные волокна более толстые, обладают мощной лактацидной анаэробной системой энергообеспечения. Они способны развивать достаточно большое усилие за короткий период времени, но быстро утомляются. Они объединяют большее количество миофибрилл. Быстрые ДЕ обеспечивают мощное быстрое (фазическое) напряжение. Среди быстрых ДЕ выделяют два подтипа: 2А - медленно утомляемый и 2В - быстро утомляемый. Эти подтипы ДЕ различаются порогом возбуждения, частотным диапазоном импульсации, а также особенностями обмена. Данные исследований физиологов, патофизиологов, морфологов показывают, что все элементы нервно-мышечной системы связаны между собой. Данная связь обеспечивается различными механизмами (аксо-нальный транспорт: медленный 1-2 мм/сут, быстрый 200-400 мм/сут; механизмы проведения нервного импульса: распад миелина при прохождении потенциала действия (ПД), синтез миелина, утилизация холина и т.п.). По данным экспертов ВОЗ (1982), периферические нервы могут рассматриваться как своеобразные аксональные кабели, отграниченныеболее или менее сложными оболочками. Эти кабели являются отростками нервных клеток и непрерывно управляются и обновляются при помощи потока молекул, что и поддерживает их морфологическую и функциональную целостность. Аксоны постоянно взаимодействуют со своими глиальными оболочками, обеспечивая метаболизм миелина. Достаточно изучено нейротрофическое влияние на мышцу, способы перестройки ДЕ при поражении различных отделов, компенсаторные механизмы нервно-мышечной системы. Обнаружен транссинаптический перенос молекул и продуктов метаболизма. При этом ортоградный ток оказывает влияние на дифференциацию мышц - доставляет пресинаптические и постсинаптические компоненты к телу клетки. Продукты распада макромолекул в теле клетки могут повторно утилизироваться при синтезе новых макромолекул. Основное назначение ретроградного транссинаптического тока -это информационный молекулярный обмен между мышцей и нейроном. Вся эта система является своеобразным сигналом для запуска метаболической перестройки нейронов, которая обеспечивает регенерацию аксонов. Таким образом, с учетом связей между мотонейроном, аксоном и мышечными волокнами, поражение одного отдела нервно-мышечной системы в дальнейшем приводит к компенсаторным (или патологическим) изменениям в других отделах. Именно эти патофизиологические особенности вызывают основные проблемы при трактовке данных комплексной ЭМГ. СТИМУЛЯЦИОННАЯ ЭЛЕКТОРОНЕЙРОМИОГРАФИЯ Стимуляционная ЭНМГ основана на анализе вызванных электрических ответов мышцы, полученных путем прямой или непрямой электрической стимуляции периферического нерва. Исследование моторного ответа мышцы и скорости распространения возбуждения по периферическим нервам Данная методика является наиболее распространенной, поскольку позволяет: 1) оценивать состояние нерва на разных его участках; 2) судить о характере поражения нерва (аксоналъное,демиелинизирующее); 3) выявлять степень поражения нерва; 4) определять состояние терминалей аксонов; 5) оценивать состояние самой мышцы. Исследование проводится с помощью накожных электродов. Кожа в месте наложения электродов обезжиривается спиртом. Активный электрод накладывают на моторную точку мышцы, референтный - на область сухожилия этой мышцы или на костный выступ, расположенный дистальнее активного электрода ("мышца-сухожилие" - "belli-tendon"). Заземляющий электрод размещается между отводящим и стимулирующим электродами. Импеданс под электродами рекомендуется установить от 5 до 10 кОм. Стимулирующий биполярный электрод накладывают в проекции нерва, иннервирующего данную мышцу, в месте наиболее поверхностного его расположения. При этом катод (-) располагают дистальнее, а анод (+) проксимальнее, так как протекающий под анодом процесс деполяризации может вызывать "анодический блок", препятствующий распространению возбуждения к мышце. Стимуляцию проводят прямоугольными импульсами обычно длительностью 0.2 мс, частотой 1 Гц, постепенно увеличивая силу тока, пока амплитуда получаемого М-ответа не перестанет нарастать. Определенная таким образом сила тока является максимальной. Для правильной оценки амплитудных и скоростных показателей используется супрамаксимальное (на 25-30% больше максимального) значение стимула. Необходимо получить стойкий по амплитуде и латентности вызванный ответ мышцы. Исследование моторного ответа мышцы М-ОТВЕТ - суммарный потенциал мышечных волокон, регистрируемый с мышцы при стимуляции иннервирующего ее нерва одиночным стимулом. В норме он представляет собой двухфазную кривую: первая фаза отрицательная (направлена вверх), вторая положительная (направлена вниз) (Рис. 2). При супрамаксимальной стимуляции в мышце гарантированно "отвечает" максимальное количество ДЕ. Поэтому М-ответ стабилен по своим параметрам, что отличает его от других волн. 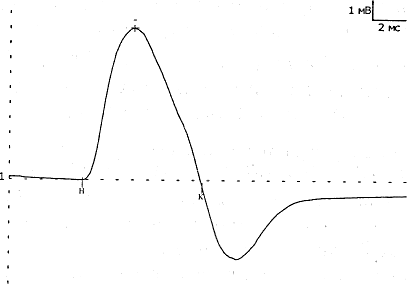 Рис. 2 М-ответ в норме. При выполнении методики надо следить, чтобы артефакт стимуляции был направлен вверх, что указывает на правильное расположение анода и катода. При снижении амплитуды М-ответа, его растянутости, полифазности необходимо проверить правильность наложения отводящих электродов, так как эти изменения имеют диагностическое значение. При исследовании гипотрофичных мышц, мышц сложной формы рекомендуется в начале установить расположение двигательной точки мышцы. Двигательная точка мышцы - участок мышцы, имеющий наименьший порог возбуждения при ее прямой стимуляции. Анатомически соответствует зоне расположения концевых пластинок терминальных ветвлений аксонов. Иногда М-ответ инвертирован. Причиной этого является либо неправильное наложение электрода (перепутаны активный и референтный электроды), либо неправильно выбранная точка стимуляции (стимулируется другой нерв), что можно видеть при стимуляции в области запястья. Причиной инверсии М-ответа может быть регистрация потенциала с соседних мышц, иннервируемых другим нервом, при наложении электродов на пораженную мышцу. Чаще всего это наблюдается при исследовании мелких мышц кисти при травмах длинных нервов. В этом случае при дистальной стимуляции М-ответ, как правило, не возникает (или крайне мал), а при стимуляции в проксимальной точке можно зарегистрировать инвертированный М-ответ от мышц, иннервируемых другим нервом. Это связано с тем, что в более проксимальных отделах длинные нервы конечностей, как правило, идут в одном сосудисто-нервном пучке (Рис. 3). 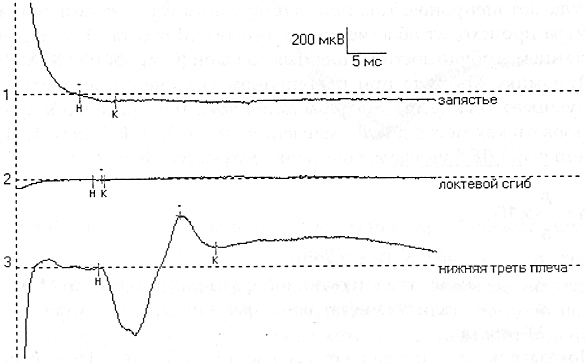 Рис. 3. Отведение от m. Abductordigit! minimi справа. Отмечается инверсия М-ответа при стимуляции в нижней трети плеча при полном анатомическом перерыве правого локтевого нерва (кривая 3). При более дистальной стимуляции (1 и 2 кривые) М-ответ не получен. При анализе результатов исследования учитываются следующие показатели: • порог раздражения - минимальная сила тока, при которой возникает М-ответ; • амплитуда негативной фазы М-ответа при стимуляции в разных точках (у длинных нервов), длительность и площадь негативной фазы; • форма М-ответа при стимуляции в разных точках; • терминальная и резидуальная латентность; • СРВм на разных участках; • динамика показателей СРВм и амплитуды М-ответа при стимуляции в разных точках по длиннику исследуемого нерва. Порог раздражения - минимальное значение стимула, способное вызвать М-ответ. Исследование проводят при усилении 100 мкВ/дел при шаге изменения стимула не более 0.1 мА. В норме порог раздражения составляет 4-6 мА (до 10 мА) при плотном прижатии стимулирующего электрода к коже. Учитывают и значение раздражителя, вызывающего максимальный М-ответ. Показано, что пороговый и надпороговый М-ответ зависят от функционального состояния пула мотонейронов. Так, при реципрокном торможении спинального центра происходит облегчение порогового М-ответа. Это связывают с нарастанием возбудимости мышечных волокон (Старобинец М.Х., 1973). Динамику М-ответа при постепенном увеличении амплитуды стимулирующего тока (с шагом нарастания тока нет более 0.5 мА) можно использовать как метод для определения числа ДЕ (McComas, 1971). Для подсчета числа ДЕ в мышце используют следующую формулу: где: n-число ДЕ в мышце,А- амплитуда негативной фазы максимального М-ответа, а-амплитуда негативной фазы примерно десяти градаций М-ответа. Показатель "а" определяют следующим образом. При усилении 100 мкВ/дел, постепенно повышая силу тока с шагом не более 0.1 мА, вызывают первые 10 дискретных уровней (ступеней) нарастания М-ответа. Определяют амплитуду негативной фазы десятой ступени. Данное значение принимается за амплитуду ответа примерно десяти ДЕ. Оценивая расчетные данные, необходимо помнить, что при стимуляции нерва потенциал отводится лишь от участка мышцы, находящегося под электродом, поэтому максимальное количество ПДЕ мы можем достоверно фиксировать только на мелких, компактных мышцах. Амплитуда М-ответа. М-ответ - достаточно стабильный потенциал при супрамаксимальной стимуляции. Считается, что негативная фаза М-ответа возникает в момент сокращения мышцы и обусловлена процессами деполяризации, позитивная фаза определяется преимущественно процессами реполяризации, которые менее синхронизированы (Рис. 4). Поэтому в настоящее время считается целесообразным проводить анализ амплитуды М-ответа по негативному пику (Гехт Б.М., Самойлов М.И., 1997). 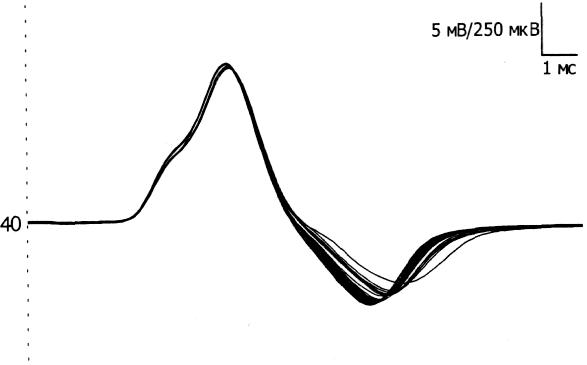 Рис. 4. Стабильность негативной фазы М-ответа и вариабельности позитивного пика при повторных стимулах. При измерении амплитуды М-ответа от изолинии до негативного пика для характеристики нормы амплитуды М-ответа введено понятие минимально допустимого значения,ниже которого определяется явная патология (Таблица 1). Показателем патологии является снижение амплитуды М-ответа при стимуляции в дистальной точке, что происходит при поражении аксонов, при мышечных процессах (как первичных, так и вторичных). Незначительно амплитуда снижается при демиелинизирующих поражениях, часто одновременно нарушается форма М-ответа, особенно при стимуляции нерва в проксимальных точках. Таблица 1. Минимально допустимая амплитуда М-ответа мышц при стимуляции в дистальной точке (измерение от изолинии до негативного пика)
Форма М-ответа. По своей природе М-ответ является результатом алгебраической суммации всех ПДЕ данной мышцы. Форма и длительность М-ответа определяются, прежде всего, синхронностью возникновения ПДЕ. В норме возникающие ПДЕ имеют некоторую асинхронность, которая связана с функциональными особенностями нерва и различной длиной терминален аксонов. Нерв состоит из большого количества нервных волокон, которые различаются скоростью проведения, что закономерно вызывает асинхронность возникновения ПД в разных ДЕ. При большем удалении точки стимуляции от мышцы степень асинхронности возрастает, что приводит к некоторому изменению формы М-ответа и в норме. Но в целом форма ответа не меняется, и при правильном наложении электродов он имеет вид двухфазной кривой, первый пик которой негативный. При демиелинизации части нервных волокон значительно увеличивается асинхронность прихода импульсов, что приводит к возникновению сильно растянутого, зазубренного М-ответа. Амплитуда М-ответа снижается незначительно. При аксональном поражении асинхронность возрастает мало, но в большей мере увеличивается амплитудная дисперсия сохранных ПДЕ, что приводит к резкому снижению амплитуды М-ответа, форма его также нарушится, но длительность уменьшится. Терминальная и резидуальная латентность. Терминальная латентность (ТЛ) - временная задержка от момента стимуляции до возникновения М-ответа при стимуляции нерва в дистальной точке. Методически ТЛ зависит от расстояния между стимулирующим и активным отводящим электродами. Поэтому для унификации исследования введено понятие резидуальной латентности (РЛ). Анатомо-физиологической основой данного понятия служит то, что нерв при входе в мышцу распадается на терминали. Терминали не имеют миелиновой оболочки, и скорость проведения импульса по ним относительно невелика. Следовательно, основную часть расстояния (от дистальной точки стимуляции до мышцы) импульс проходит по миелинизированному волокну и лишь небольшую - внутри мышцы по немиелинизированной терминали. Поэтому, если из ТЛ вычесть то время, за которое импульс проходит расстояние от точки стимуляции до мышцы, то полученная разность, называемая резидуальной латентностью (РЛ), будет отражать время прохождения импульса по терминалям аксонов. Время прохождения импульса по миелинизированной части рассчитывается как S/V, где S - терминальное расстояние, измеряемое от активного отводящего электрода до катода стимулирующего электрода, V - скорость проведения импульса в дистальном сегменте данного нерва. РЛ = ТЛ-S/V РЛ значительно возрастает при терминальных полиневропатиях, когда процесс начинается с синапсов и терминален, при токсических поражениях. По данным литературы, рекомендуется для нормирования РЛ использовать максимальное допустимое значение РЛ,которое для периферических нервов рук составляет 2.5 мс, для ног - 3.0 мс. С учетом некоторого несоответствия указанных нормативных данных получаемым на практике значениям мы проанализировали 221 срединный нерв и 222 локтевых нерва. Оказалось, что по локтевому нерву РЛ составила 1.8±0.55 мс (1.25-2.35), по срединному нерву 2.410.59 мс (1.81-2.99). Как видно, по срединному нерву РЛ оказалась выше. Такое различие между локтевым и срединным нервом определяется особенностями измерения расстояния по срединному нерву. Для получения сравнимого с нормативами результата расстояние измеряется по ломаной линии от катода стимулирующего электрода до головки многогранной кости, далее вдоль Thenar до активного электрода. Исходя из проведенного исследования и указанных выше особенностей измерения расстояния, в качестве максимально допустимого значения РЛ для срединного нерва можно принять значение – 3 мс. Расчет резидуальной латентности возможен только при анализе длинных нервов, которые можно стимулировать более чем в одной точке. При анализе проведения по коротким нервам сохраняет свое значение понятие терминальной латентности. К коротким можно отнести нервы, которые доступны для стимуляции только в одной точке (например, п. axillaris). При исследовании мышц лица измеряется только ТЛ. В норме она не должна быть выше 3.5-4 мс. При исследовании коротких нервов большое методическое значение имеет точное симметричное наложение электродов и соблюдение основных нормативных расстояний между стимулирующим и отводящим электродами. Для адекватного применения нормативов латентности важно правильное измерение расстояния от катода до активного электрода. |