Учебное пособие для ординаторов по специальности Функциональная диагностика
 Скачать 2.86 Mb. Скачать 2.86 Mb.
|

|
Наиболее часто исследуемые нервы. Срединный нерв. Для определения локализации компрессии срединный нерв условно разделяют на пять участков соответственно гаги доступным для прямой стимуляции точкам: самая проксимальная точка — точка Эрба находится в области надключичной ямки, следующая — в области подмышечной впадины, далее — на плече, вышелоктя, следующая — на предплечье ниже локтя, и, наконец, на запястье. Таким образом, скорость можно определить на четырех участках; величину СП на участке от запястья до thenar оценивают по величине резидуальной латенции РЛ. На представлены также М-ответы, вызванные стимуляцией всех этих точек. Синдром карпального канала. При прохождении самого дистального участка срединного нерва через запястье он может подвергаться сдавлению в карпальном канале. При электродиагностике все величины СП по разным участкам остаются в пределах нормы. Может быть снижена лишь СП по дистальному участку. Значительное снижение СП наблюдается дистальнее запястья, величина РЛ может достигать 4—6 мс (при верхней границе нормы 2,5 мс). Основное место в диагностике этого синдрома принадлежит ишемической пробе, так как только у этих больных ее применение вызывает еще большее увеличение РЛ. Компрессия срединного нерва связкой Пирогова выше локтя приводит к локальному снижению СП через область локтевого сгиба, СП по другим участкам может быть в пределах нормы. Может, однако, отмечаться небольшое снижение СП по участку дистальнее места компрессии, увеличение РЛ и снижение амплитуды М-ответа. Ишемия вызывает ненормально сильное падение СП только в области компрессии. Синдром пронатора — компрессия срединного нерва ниже локтевого сустава m. pronatortheres. СП по всем участкам сохраняется в пределах нормы, в том числе через область локтевого сгиба. СП по дистальному участку снижена, РЛ может быть увеличена, амплитуда М-ответа снижена. Ишемия приводит к очень сильному падению СП только на участке ниже локтевого сгиба и запястья. Синдром выходного отверстия — компрессия срединного нерва в области ключицы. СП снижена только на участке между точкой Эрба и подмышечной впадиной. В дистальных участках СП остается нормальной. Остальные параметры остаются в норме, но может произойти извращение СП, т. е. СП по более проксимальным отделам могут быть ниже, чем по более дистальным. Ишемическую пробу проводить нецелесообразно. Для определения места локализации поражения локтевого нерва последний условно разделяется также на несколько участков (рис. 13). 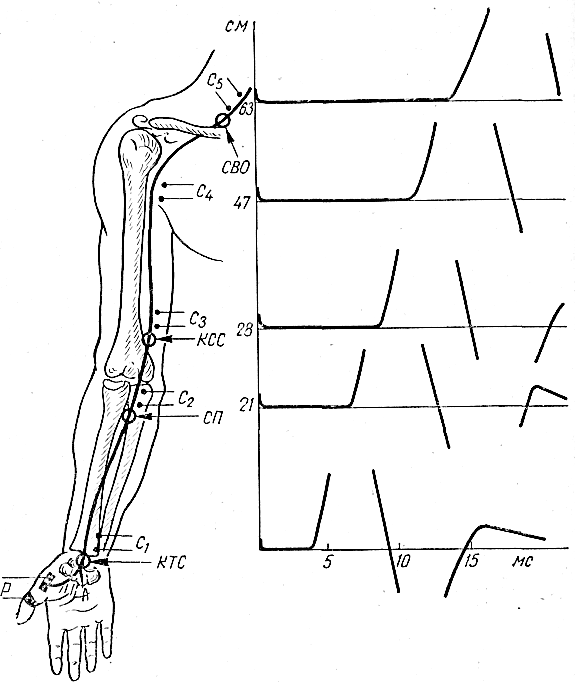 Рис. 13.Схема расположения срединного нерва, точки стимуляции и ПД, зарегистрированных при супрамаксимальной стимуляции срединного нерва соответственно в точках Q, С2, С3, Cj, Cs и отведении ПД приводящей мышцы большого пальца. Р — расположение отводящих электродов; КТС — место возможной компрессии нерва при карпальном туннельном синдроме; СП — то же при синдроме пронатора; КСС — то же при компрессии связкой Пирогова; СВО — то же при синдроме выходного отверстия. На оси абсцисс миллисекунды, на оси ординат — расстояния от точки d до точек С2, Сз, С, и С5. Нормальные значения: Амп.(мВ) >3,5 СРВ(м/с) >50 Рез.Лат (мс) <2,5 Локтевой нерв. Кубитальный туннельный синдром — компрессия локтевого нерва в локтевом туннеле. Величины СП на участке подмышечная впадина — выше локтя и от точки ниже локтя до запястья могут быть в пределах нормы -60 м/с, СП через локоть (короткий сегмент) снижена 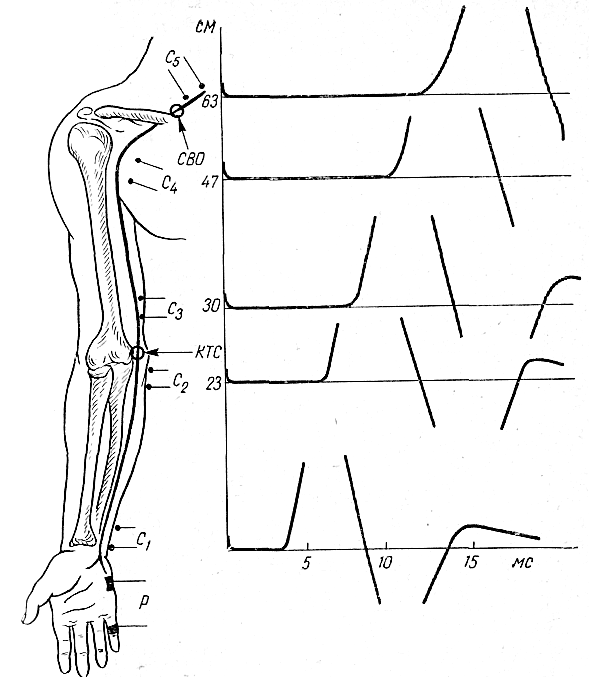 Рис. 14.Схема расположения локтевого нерва, точек стимуляции и ПД, зарегистрированных при супрамаксимальной стимуляции локтевого нерва соответственно в точках Сь С2, С3, С4 и gs и отведении ПД отводящей мышцы мизинца. Р — расположение отводящих электродов; КТС — место возможной компрессии нерва при кубитальном туннельном синдроме; СВО — то же при синдроме выходного отверстия. На графиках представлены те же величины, что и для срединного нерва на рис. 16. До 15 м/с (W.Bradley, 1974). СП по проксимальному участку равна СП по дистальному, что не наблюдается в норме. Ишемическая проба вызывает чрезмерное падение СП на коротком сегменте. РЛ в пределах нормы или увеличена, амплитуда М-ответа может быть снижена. На нижних конечностях компрессии подвергаются главным образом малоберцовый и большеберцовый нервы. Нормальные значения: Амп.(мВ) >3,5 СРВ(м/с) >50 Рез.Лат (мс) <2,5 Малоберцовый нерв. Р 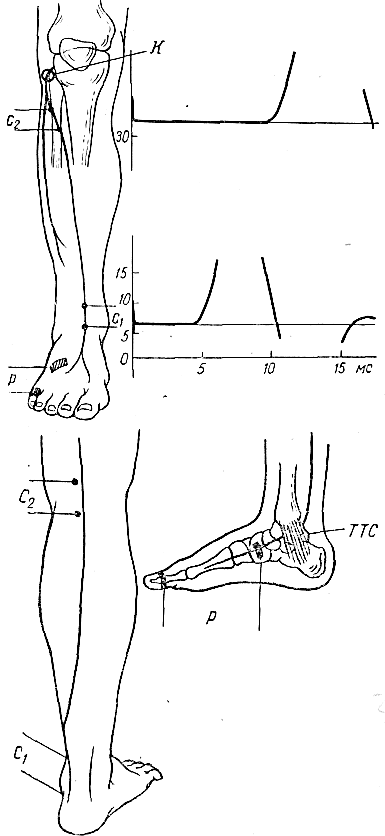 ис. 15.Схема "расположения малоберцового нерва, точек стимуляции и ПД, зарегистрированных при супрамаксимальной стимуляции малоберцового нерва в точках С1 и С2 и отведении ПД короткого разгибателя пальцев. ис. 15.Схема "расположения малоберцового нерва, точек стимуляции и ПД, зарегистрированных при супрамаксимальной стимуляции малоберцового нерва в точках С1 и С2 и отведении ПД короткого разгибателя пальцев.Р — расположение отводящих электродов; К — место возможной компрессии нерва. Нормальные значения: Амп.(мВ) >3,0 СРВ(м/с) >40 Рез.Лат (мс) <3,5 Большеберцовый нерв. 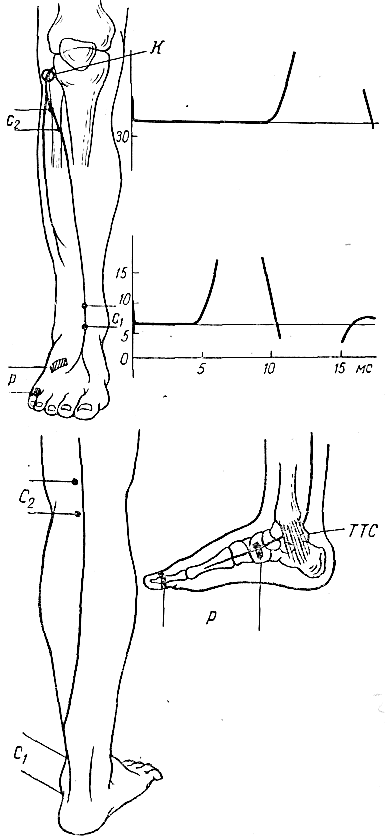 Рис. 16.Схема расположения большеберцового нерва и точек стимуляции С1, C2. Р — расположение отводящих электродов при отведении ПД короткого сгибателя большого пальца; ТТС— место возможной компрессии нерва при тарзальном туннельном синдроме. Нормальные значения: Амп.(мВ) >3,0 СРВ (м/с) >40 Рез.Лат (мс) <3,5 Синдром выходного отверстия диагностируется так же, как синдром выходного отверстия срединного нерва. Компрессия большеберцового нерва возникает чаще непосредственно перед разделением его на поверхностную и глубокую ветви в области головки малоберцовой кости. Точки стимуляции — в подколенной ямке и в области медиальной лодыжки. Тарзальный туннельный синдром обусловлен компрессией большеберцового нерва в самом дистальномего участке-месте прохождения в фасциальном туннеле на уровне медиальной лодыжки. Характерное электрофизиологическое проявление его — увеличение резидуальной латенции. Исследование скорости проведения по чувствительным нервам также находит широкое применение в неврологической клинике. Наиболее адекватным методом, позволяющим оценить степень повреждения чувствительных волокон, является метод, предложенный Dawson (1956), lile, Sera (1958), основанный на записи нейрограммы по ходу распространения импульса при раздражении чувствительных волокон кончиков пальцев с помощью кольцевых электродов. При таком методе возможна регистрация ПД нерва, состоящего лишь из чувствительных волокон по всему его длиннику. Это дает возможность дифференцированно оценить скорость проведения на каждом участке нерва, избежать недостатков, связанных с включением в ответ и части двигательных волокон в случае антидромного и ортодромного (при раздражении в более проксимальных его участках) раздражения нерва. Оцениваемыми параметрами являются: сила тока, при которой возникает ответ (в норме она находится в границах 5 мА), амплитуда вызванного ПД нерва (8— 45 мкВ для верхних конечностей; 12—15 мкВ—для нижних); скорость проведения (в дистальных отделах нервов конечностей она составляет 65 м/с, в проксимальных— на 10 м/с больше) и латентный период ответа. В патологических состояниях при различных формах невропатий (диабетической, алкогольной и др.) могут отмечаться значительные изменения этих параметров:резкое уменьшение амплитуды ПД нерва (вплоть до полного отсутствия ПД нерва) при значительном увеличении силы тока, уменьшение скорости проведения по нервам и удлинение латентных периодов. Исследование; скорости проведения возбуждения по чувствительным нервам в связи с малой амплитудой ПД возможно только при наличии специальных приборов (усреднителей), позволяющих выделить и зарегистрировать ПД чувствительного нерва. ЭЛЕКТРОМИОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА СОСТОЯНИЯ НЕРВНО-МЫШЕЧНОЙ ПЕРЕДАЧИ Определение состояния нервно-мышечной передачи лежит в основе диагностики всех патологических процессов, обусловленных нарушениями передаточной функции синапсов, т. е. при миастении и всех миастенических синдромах, ряде интоксикаций, а также для определения глубины релаксации мышц в анестезиологической практике. Методы современной клинической ЭМГ позволяют не только определить степень надежности нервно-мышечной передачи, но и характер нарушений функции синапса, тип блока и механизмы его развития [Гехт Б. М. и др., 1974; Самойлов М. И., 1976; Slomicetal. 1968;EmericВ., 1971] и др.. Наиболее широкое распространение в настоящее время получили два метода изучения нервно-мышечной передачи — использование стимуляционной электромиографии и регистрация потенциалов действия одиночных мышечных волокон — изучение «джиггер-феномена». АНАЛИЗ ПАРАМЕТРОВ М-ОТВЕТА М-ответ — ВП мышцы, являющийся суммарным синхронным разрядом ДЕ мышцы в ответ на электрическое раздражение нерва. Обычно М-ответ регистрируют с помощью поверхностных (накожных) электродов, которые более объективно и полно отражают суммарную активность мышцы по сравнению с игольчатыми электродами. При изучении М-ответа обращают внимание на интенсивность порогового раздражения, латентный период ВП, его форму, амплитуду, длительность, а также на взаимозависимость этих показателей. Необходимо регистрировать порог раздражения (порог М-ответа) — минимальное электрическое раздражение, способное вызвать М-ответ. Величина пороговой стимуляции в обычных условиях может варьировать в небольших пределах в зависимости от характера наложения электродов, индивидуальных особенностей кожного сопротивления (сухость, влажность, сальность кожных покровов), достаточной влажности марлевых подэлектродных прокладок. Поэтому чрезвычайно важное значение имеет подготовка к исследованию — обработка кожи в месте наложения электродов спиртом, приготовление свежих подэлектродных прокладок и достаточное смачивание их (чрезмерное смачивание кожи, как и недостаточное, может вызвать артефакты записи). Грубое повышение порога М-ответа наблюдается при поражении нерва или мышцы, снижение порога встречается крайне редко. Определяют также интенсивность раздражения, вызывающего М-ответ максимальной амплитуды (максимальный М-ответ). Амплитуда М-ответа при максимальном раздражении может иметь некоторые колебания ввеличине, что определяется непостоянным включением наиболее высокопороговых ДЕ. С целью гарантированного охвата всех функционирующих ДЕ мышцы используют так называемое супрамаксимальное раздражение — повышение силы раздражения после-достижения максимального М-ответа еще на 25—50%. Форма М-ответа зависит от ряда факторов. При биполярной отведении М-ответ имеет негативную и позитивную фазы соответственно прохождению волны возбуждения под обеими электродными пластинами. Этим обусловлено оптимальное расстояние между пластинами — 15—20 мм, при котором обе фазы потенциала непрерывны и не накладываются друг на друга. При описании формы ПД мышцы его характеризуют как монофазный (при отклонении от изолинии в одну сторону), двухфазный (при отклонении от изолинии последовательно в одну, а затем в другую сторону), трех-, четырех- или полифазный. В норме на форму М-ответа наибольшее влияние оказывает положение отводящих электродов. Как уже было отмечено, основной электрод должен размещаться на брюшке исследуемой мышцы перпендикулярно ходу мышечных волокон, а индифферентный — на сухожилии. При перемещении брюшкового электрода форма М-ответа существенно изменяется. Этим определяется необходимость до начала исследования подобрать такое положение отводящего электрода относительно мышцы, при котором форма мышечного потенциала была бы правильной двухфазной. Лишь при невозможности получить двухфазный М-ответ следует рассматривать его как трех-, четырех- или полифазный. Изменение формы мышечного потенциала может выявляться также при патологии части нервных или мышечных волокон как проявление избирательного выключения некоторых ДЕ. 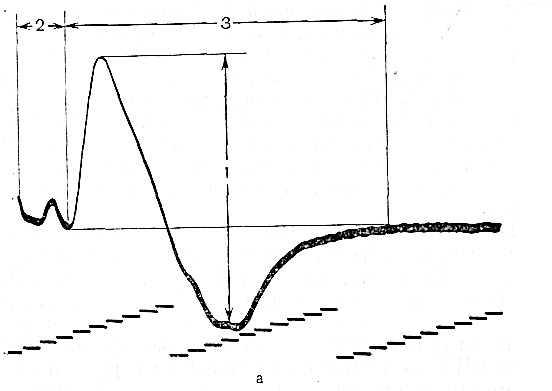 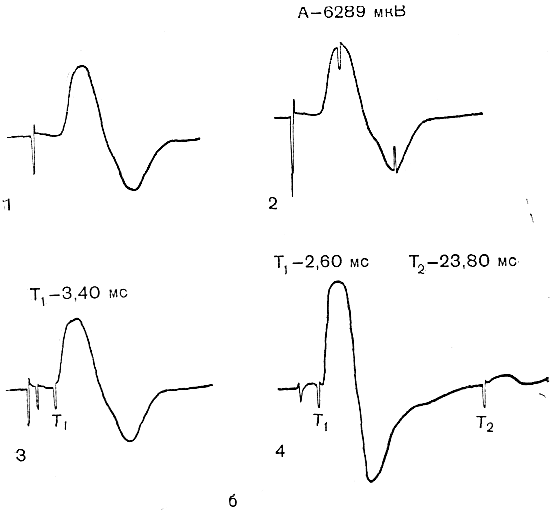 Рис. 17. М-ответ: а — амплитуда (1), латентность (2), длительность (3) М-ответа; б — М-ответ на аппарате «Меделек МС-92»: 1 — общий вид; 2—4— автоматический подсчет амплитуды (2), латентности (3), длительности (4). Амплитуду М-ответа определяют как от негативного до позитивного пика, так и от негативного пика до изолинии. Обычно используют первый способ. Максимальная амплитуда М-ответа, получаемая при супрамаксимальной стимуляции, отражает суммарный ответ ДЕ и, следовательно, их число в исследуемой мышце. В этом отношении результаты, получаемые с помощью игольчатых электродов, значительно менее надежны по сравнению с методикой использования поверхностных электродов. F. Buchthal (1955), отмечая недостатки отведения М-ответа игольчатыми электродами, указывает на невозможность оценить с помощью игольчатых электродов число мышечных волокон, отвечающих на стимуляцию нерва, и считает этот метод необъективным для количественного измерения прогрессирующей дегенерации мышечных волокон, для локализации блока невральной проводимости и тестирования нервно-мышечной передачи. Амплитуда максимально М-ответа измеряется в милливольтах или микровольтах и, как считали большинство авторов, имеет значительные индивидуальныe различия. На этом основании Ст. Байкушев с соавт. (1974) и некоторые другие исследователи считали диагностическую эффективность измерения амплитуды М-ответа невысокой. Следует, однако, отметить, что индивидуальная вариабельность величины амплитуды М-ответа в значительной степени определяется небрежным, нестандартным наложением отводящих электродов над мышцей и нарушением других технических требований методики. Во всяком случае, скепсис в отношении возможности анализа максимальной амплитуды М-ответа в диагностических целях серьезно поколеблен в последнее время в связи с исследованием изменения амплитуды М-ответа при поражении центральной и периферической соматической нервной системы, а так же структур вегетативной нервной системы. Широкое распространение получил метод анализа соотношения между максимальной и минимальной амплитудой М-ответа. При некоторых состояниях (физиологических и патологических) определяется диссоциация между максимальным и минимальным (пороговым) М-ответом: амплитуда максимального М-ответа снижается, а амплитуда порогового повышается, и наоборот. Длительность М-ответа (максимального по амплитуде) определяется в миллисекундах как время прохождения импульса с момента первого отклонения от изолинии до момента возврата к изолинии в конце его. Поскольку в конце вызванного мышечного ответа миографическая кривая нередко долго сохраняет некоторые колебания (что затрудняет объективную фиксацию момента возврата к изолинии), Н. Е. Kaeser (1975) и др. предлагают вычислять продолжительность только негативной фазы (от начала потенциала до начала позитивной фазы), однако, учитывая различную продолжительность негативной и позитивной фаз, следует, по-видимому, отдать предпочтение первому способу. Длительность М-ответа определяют обычно при стимуляции нерва в возможно дистальной точке (в области запястья для срединного и локтевого нервов и в области голеностопного сустава при стимуляции нервов голеней конечности), так как при стимуляции в более проксимальных отделах нерва форма, амплитуда и длительность М-ответа, как правило, изменяются. Длительность М-ответа измеряют как разность времени возвращения кривой к изолинии после М-ответа, т.е. от артефакта раздражения до конца мышечного потенциала латентного периода М-ответа (время от артефакта раздражения первого отклонения от изолинии в начале ВП). Для правильного определения длительности М-ответа имеет значение стабильноерасстояние между электродными пластинами, правильность их размещения над мышцей, а также и от самой мышцы (ее размеров и функционального назначения). Последние исследования показали также зависимость длительности М-ответа от состояния интерстициальных соединительнотканных прослоек в мышечной ткани (Скворцов И. А. и др., 19811), а также от патологии двигательных волокон нервного ствола, вовлечения эндоневрия (паренхиматозные или интерстициальные полиневропатии). Анализ продолжительности латентного периода М-ответа прежде всего используется для определения СПИ по двигательным волокнам иннервирующего мышцу нерва. В меньшей степени этим показателем можно характеризовать сам ПД мышцы. Поэтому методика измерения латентности М-ответа и анализ факторов, влияющих на нее, представлены ниже в разделе, посвященном определению скорости моторного проведения по нерву. МЕТОДИКА ОПРЕДЕЛЕНИЯ F-ОТВЕТА. F-ответ – поздний ответ, периодически регистрируемый с мышцы при антидромной активации мотонейронов. Впервые F-ответ описан MagladeryandMcDougal (1950). F-ответ вызывается при супрамаксимальной электрической стимуляции периферического нерва (J. Kimura, 1974, C.P.Panayiotopoulosetal., 1977). Физиологическая характеристика метода и клиническое значение параметров F-волны при патологии При неизменных условиях стимуляции и регистрации F-волны ее амплитуда, латентность, длительность и форма меняются при каждом последующем стимуле, что обусловлено индивидуальными особенностями антидромной активации разных мотонейронов передних рогов спинного мозга от стимула к стимулу. F-волна представляет собой активность одного мотонейрона или суммарную активность нескольких мотонейронов. F-ответ составляет активность всего около 1-5% числа мотонейронов, участвующих в формировании М-ответа. Схема формирования F-волны представлена на рисунке. Супрамаксимальная стимуляция нерва приводит к ортодромному распространению стимула в дистальном направлении и активации всей мышцы с формированием М-ответа, который регистрируется с латентным периодом 3-15 мс. Одновременно с ортодромным распространением импульса при стимуляции нерва возникает и антидромно направленная активация, которая охватывает тела всех мотонейронов. Возникающая вслед за этим реактивация охватывает только наиболее возбудимые в этот момент мотонейроны. Потенциал действия от возбужденных клеток мотонейронов распространяется уже ортодромно к мышце, потенциал которой обозначается F-волной и регистрируется с несколько большим латентным периодом, чем зарегистрированный М-ответ (20-40 мс). При последующих раздражающих стимулах происходит реактивация других мотонейронов, являющихся наиболее возбудимыми в момент активации. Разное количество реактивированных мотонейронов и разные свойства проводимости по соответствующим аксонам определяют разную амплитуду, длительность, латентность, связанную с ней скорость и площадь F-волны (рис18). Показатели латентности 10-40 F-волн, зарегистрированные у одного испытуемого, колеблются в определенных пределах, характеризуя состояние проводимости по нервным волокнам от максимальной скорости проведения до минимальной. Для оценки этого состояния нервных проводников используются показатели минимальной латентности, максимальной латентности, средней латентности и показатель разброса латентности. Соответственно рассчитываются показатели минимальной, максимальной, средней СПИ и тахеодисперсии. 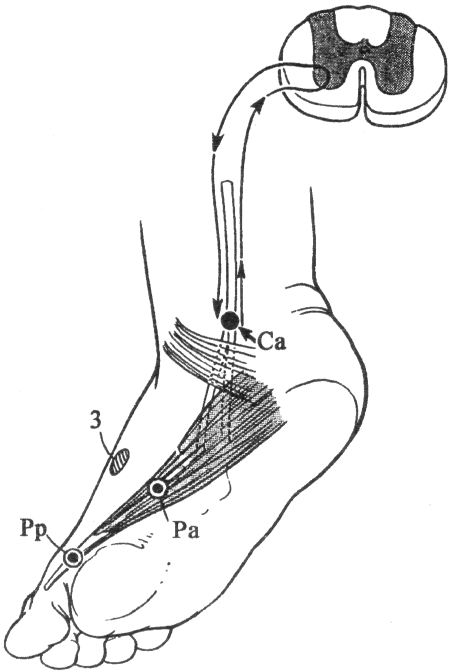 Рис.18.Схема регистрации F-волны при стимуляции большеберцового нерва (По J.A.Liveson, 1992). Ра - регистрирующий активный электрод, Рр - регистрирующий референтный электрод, Са-стимулирующий активный электрод, 3 — заземляющий электрод. |