все лекции по биологии. Учебное пособие для студентов I курса, обучающихся по специальностям лечебное дело и медикодиагностическое дело
 Скачать 1.56 Mb. Скачать 1.56 Mb.
|

|
Тема № 2. Хромосомный и геномный уровни организации наследственного материала. Морфофизиологическая характеристика метафазной хромосомы. Типы и правила хромосом. Хромосомный уровень организации наследственного материала характеризуется особенностями морфологии и функций хромосом. Роль хромосом в передаче наследственной информации была доказана благодаря: 1) открытию хромосомного определения пола; 2) установлению групп сцепления генов; 3) построению генетических и цитологических карт хромосом. Метафазная хромосома (спирализованный хроматин) состоит из двух хроматид. Форма определяется наличием первичной перетяжки – центромеры. Она разделяет хромосому на 2 плеча. Расположение центромеры определяет основные формы хромосом: метацентрические (равноплечие), субметацентрические (неравноплечие), акроцентрические (резко неравноплечие) и телоцентрические (одноплечие). Степень спирализации хромосом неодинаковая. Участки хромосом со слабой спирализацией называют эухроматиновыми. Это зона высокой метаболической активности, где ДНК состоит из уникальных последовательностей. Зона с сильной спирализацией – гетерохроматиновыйучасток, обеспечивающий транскрипцию. Различают конститутивный гетерохроматин – генетически инертный, несодержий генов и непереходящий в эухроматин, а так же факультативный, который может переходить в активный эухроматин. Концевые отделы дистальных участков хромосом называют теломерами. Теломеры обеспечивают стабильность структуры хромосом и ограничивают число клеточных делений. Хромосомы обладают следующими свойствами (правилами хромосом): 1. Индивидуальности – негомологичные хромосомы различаются между собой. 2. Парности – гомологичные хромосомы образуют пары. 3. Постоянством числа – все соматические клетки организма имеют постоянное число хромосом, характерное для вида. 4. Непрерывности – репродукция хромосом и передача наследственной информации при делении клетки. Кариотип и идиограмма. Характеристика кариотипа человека. Хромосомы подразделяются на аутосомы и половые хромосомы или гетерохромосомы. По предложению Г.А. Левитского (1924) совокупность признаков диплоидного набора хромосом соматических клеток была названа кариотипом. Он характеризуется числом, формой и размерами хромосом. В кариотипе соматической клетки человека различают 22 пары аутосом и пару половых хромосом. Набор хромосом в соматических клетках называют диплоидным, а в половых клетках – гаплоидным. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы – систематизированного расположения хромосом попарно в проядке убывания их величины. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и форме. По Денверской классификации хромосомы человека делят на 7 групп, которые обозначают буквами латинского алфавита: A: 1-3 – крупные метацентрические. B: 4-5 – крупные субметацентрические. C: 6-12 и Х-хромосома – средние метацентрические и субметацентрические. D: 13-15 – средние акроцентрические. E: 16-18 – малые субметацентрические. F: 19-20 – малые метацентрические. G: 21-22 и Y-хромосома – малые акроцентрические. Согласно Парижской классификации (1971 г.) хромосомы различают по их дифференциальной окраске. Расположение окрашенных участков по длине хромосомы зависит от локализации определенных фракций ДНК. Характер окрашивания специфичен для негомологичных хромосом. Молекулярная организация хромосом эукариот. Хромосомы эукариот – это спирализованный хроматин – комплекс ДНК и белков, где 40% приходится на ДНК, 40% – на гистоновые (основные) белки и почти 20% – на негистоновые белки и немного РНК. Гистоны – хромосомные белки с высоким содержанием аргинина и лизина. Их пять классов: H1, H2A, H2B, H3, H4. Гистоны стабилизируют структуру хромосомы и играют роль в регуляции активности генов. Негистоновые (кислые) белки. В хромосомах их количество приблизительно вдвое меньше гистоновых. Существует более 100 видов негистоновых белков. Они разнообразны по молекулярному весу, структуре, видоспецифичны. Эти белки могут быть ответственны за репликацию, репарацию, транскрипцию, возможно, играют роль и в активации генов. К ним относят актин, миозин, тубулин, ферменты синтеза РНК и ДНК полимеразы и другие. Из пяти классов гистонов, четыре (Н2А, Н2В, Н3, Н4) образуют своеобразные шаровидные тельца – коры диаметром около 10 нм. В одну кору входит 8 молекул гистонов. Отрезок двуспиральной нити ДНК (около 140 нуклеотидных пар) образует вокруг нее почти два оборота. Кора, вместе с молекулой ДНК, образует уникальную повторяющуюся единицу организации наследственного материала эукариот – нуклеосому. Соседние нуклеосомы соединены друг с другом короткими линкерными отрезками ДНК (1 10 нм или 30-100 пар нуклеотидов), что формирует хроматиновую или нуклеосомную нить. К каждому такому отрезку присоединены молекулы гистона Н1. Допускается, что вследствие взаимодействия Н1 с нуклеосомами происходит конденсация хроматиновой нити (d=10 нм), что формирует хроматиновую спираль (d=25нм). Уровни упаковки генетического материала. Таким образом, уровни упаковки ДНК следующие: 1) Нуклеосомная нить; 2) Супернуклеосомный – хроматиновая спираль; 3) Хромонемный – уложенная петлями и спирализованная хроматиновая спираль. 4) Хромосомный – четвертая степень спирализации ДНК. На этом уровне уложенная петлями хроматиновая спираль спирализуется еще раз и формирует хроматиду, которая является структурным элементом хромосомы. В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок называется эухроматином (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными. Геномный уровень организации наследственного материала. Геном – совокупность всех генов гаплоидного набора хромосом данного вида организма. Геномный уровень организации наследственного материала имеет особенности у прокариот и эукариот. В геноме бактерий подавляющее большинство генов уникальны. Исключением являются гены, кодирующие р-РНК и и-РНК. Эти гены повторяются в геноме бактерий несколько раз. Следует отметить определенное несоответствие между числом пар нуклеотидов в геноме бактерий и числом генов в них. Так, ДНК кишечной палочки содержит 4,6 млн. пар нуклеотидов. Структурных генов у них около 1000, на которые приходится 1 1,5 млн. пар нуклеотидов. Остается предположить, что значительную часть в ДНК бактерий составляют участки, функции которых пока не ясны. Геном эукариот характеризуется:
Для эукариот также характерна избыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года около 31 тыс. генов. Больше половины генома эукариот составляют уникальные гены, представленные лишь по одному разу. У человека таких уникальных генов – 64% от количества всех структурных генов, у теленка – 55%, у дрозофилы – 70%. Морган указал на стабильность структуры генома и постоянство расположения генов в хромосомах. В 70-х годах у дрозофилы обнаружена группа генов, представленных многими кочующими генами, которые разбросаны по разным участкам хромосом. Во многих геномах они находятся в изобилии: например, кочующие гены составляют до 50% человеческой ДНК. Сформировалось представление, что в состав генома про- и эукариот входят гены:
Генетическая система клетки: ядерные гены и плазмогены. Хромосомная теория наследственности указывает на ведущую роль ядра в передаче наследственных признаков. По мере развития генетики накапливались данные о возможности прямого участия в явлениях наследственности цитоплазмы. Такая форма наследственности определяется органоидами клетки, способными к самовоспроизведению, благодаря наличию в них ДНК: митохондриями и пластидами. Наследственность, при которой материальной основой наследования являются элементы цитоплазмы, называется цитоплазматической. Такая форма наследственности не подчиняется законам Менделя. Цитоплазматическое наследование, в отличие от хромосомного, осуществляется по материнской линии, т.к. цитоплазмой богата яйцеклетка, а не сперматозоид. Наследственные факторы цитоплазмы и органелл называются плазмотип или плазмон. Единица цитоплазматической наследственности – плазмоген. У прокариот носителями цитоплазматической наследственности является ДНК плазмид. Плазмотип эукариот состоит из наследственного аппарата пластид и митохондрий. Цитоплазматические наследственные структуры распределяются неравномерно в дочерние клетки, в отличие от ядерных ДНК. Среди типов цитоплазматической наследственности выделяют: I. Собственно цитоплазматическое наследование.
II. Предетерминация цитоплазмы. III. Наследование через инфекции. Пластидная наследственность. Наследование пестролистности у растений является примером такого типа. Пестролистность у цветка ночной красавицы наблюдается только у женского растения. Признак пестролистности связан с мутациями ДНК в хлоропластах, что приводит к их обесцвечиванию. Митохондриальная наследственность. Митохондриальные гены кодируют 2 группы признаков, связанных:
В митохондриях дрожжевых клеток обнаружены гены дыхательных ферментов. Эти гены находятся в плазмидах. У бактерий выделяют 3 типа плазмид:
Бактерии с фактором F являются мужскими. При конъюгации фактор F переходит в женскую особь и она становится мужской. Фактор R обеспечивает устойчивость к антибиотикам и также передается при конъюгации. Фактор col содержится в плазмидах, ДНК которых кодирует белки-колицины, убивающие бактерии, не содержащие таких плазмид. Цитоплазматическая мужская стерильность (ЦМС) – разновидность внеядерной наследственности. Характеризуется присутствием в ДНК митохондрий и пластид цитоплазматического гена (плазмогена), угнетающего формирование хромосом при образовании пыльцы. В результате появляется нежизнеспособная (стерильная) пыльца, не образующая спермиев. ЦМС играет большую роль в селекции и семеноводстве для исключения самоопыления и последующего получения гетерозисных гибридов (кукуруза, лук, пшеница, свекла и др.) Предетерминация цитоплазмы. В цитоплазме яйцеклетки присутствует негенетические факторы (ферменты, белки-репрессоры), влияющие на экспрессию ядерных генов. Возникает "материнский эффект" – влияние генотипа матери на развитие потомства, передаваемое через цитоплазму яйцеклетки. Так наследуется направление закручивания спирали раковины прудовика, детерминируемое генотипом матери, а не зародыша. Материальным субстратом цитоплазматической наследственности выступают гены ядерной ДНК, пластид, митохондрий и какие-то пока не установленные факторы. Система генетического аппарата клетки включает генотип ядра и плазмотип (плазмон) цитоплазмы (Рис. 2). Генетический аппарат клетки дискретен. В генотипе ядра он представлен хромосомами и генами, входящими в состав хромосом, в плазмотипе цитоплазмы – плазмогенами, которые являются фрагментами пластидной и митохондриальной ДНК. 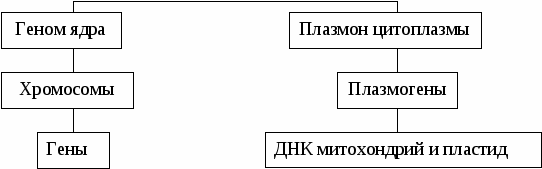 Рис. 2 — Система генетического аппарата клетки Тема № 3. Экспрессия генов у про- и эукариот. Общее понятие генетического материала и его свойства. Роль ДНК в передаче наследственной информации была доказана с открытием явлений трансформации, трансдукции и конъюгации у микроорганизмов. Конъюгацией – процесс, при котором осуществляется перенос генетической информации. Трансформация – это способность одного штамма бактерий встраивать участки молекулы ДНК другого штамма и приобретать при этом свойства последнего (опыты Ф. Гриффита с вирулентными и невирулентными штаммами пневмококков). Трансдукция – это способность бактериофагов переносить фрагменты ДНК от одного штамма бактерий к другому и передавать соответствующие свойства. Генетический материал – компоненты клетки, структурно-функциональное единство которых обеспечивает хранение, реализацию и передачу наследственной информации при вегетативном и половом размножении. Генетический материал обладает универсальными свойствами живого: дискретностью, непрерывностью, линейностью, относительной стабильностью. Основными свойствами генетического материала являются:
- диплоидным набором хромосом; - двойной спиралью ДНК; - вырожденностью генетического кода; - повтором некоторых генов; - репарацией нарушенной структуры ДНК. Строение гена у про- и эукариот. Первоначально предполагалось, что ген является неделимой, целостной единицей. В целом ген подвергается мутациям, рекомбинациям и выполняет основную функцию – кодирование белка. Однако оказалось, что ген дискретен. Наиболее четко дискретность гена была изучена американским генетиком С. Бензером на примере исследований тонкой структуры генов фага Т4 кишечной палочки. Им было показано, что ген может быть разделен кроссинговером на множество частей. Дискретная организация генов была установлена и у эукариот. В конце 50-х годов Бензер предполагал, что ген одновременно является целостной и дискретной единицей. При выполнении основной функции – программировании синтеза белка – ген выступает как целостная единица, изменение которой вызывает изменение структуры белковой молекулы. Эту единицу Бензер назвал цистроном. По величине он примерно равен гену. Дискретность гена заключается в наличии субъединиц. Элементарная единица изменчивости, единица мутации названа мутоном, а единица рекомбинации – реконом. Минимальные размеры мутона и рекона равны 1 паре нуклеотидов и называются сайт. Таким образом, сайт – это структурная единица гена. Кодон – функциональная единица гена (единица генетического кода, три нуклеотидных остатка (триплет) в ДНК или РНК, обычно кодирующих включение одной аминокислоты в полипептид). Размеры генов различны. Средний размер гена у человека составляет около 27 тысяч пар нуклеотидов. Самые короткие гены содержат всего два десятка нуклеотидов, например, гены эндорфинов – белков, вызывающих ощущение удовольствия. Гены интерферонов – белков, защищающих человека от вирусных инфекций, имеют размер около 700 пар нуклеотидов. Самый длинный ген, кодирующий один из белков мышц – миодистрофин, содержит 2,5 миллиона пар нуклеотидов. Вплоть до конца 70-х годов полагали, что гены существуют в виде целого отрезка ДНК. Однако в 1977 г. было показано, что у аденовируса некоторые гены существуют не в виде целого отрезка ДНК, а в виде фрагментов, распределенных вдоль генома. Последовательность нуклеотидов, составляющая мозаичный ген, вначале переписывается в молекулу про-и-РНК, которая является своего рода предшественником и-РНК Участки, несущие информацию, названы экзонами, а не содержащие ее – интронами. Например, ген цепи β-глобулина человека включает в себя 3 экзона и 2 интрона: ген постоянного участка тяжелой цепи иммуноглобулинов мыши содержит 4 экзона и 4 интрона. Затем про-и-РНК подвергается процессингу и сплайсингу. После этого получается и-РНК, готовая для последующей транскрипции. Окончательного объяснения факта существования интронов пока не найдено. Допускается, что в момент образования и-РНК из про-и-РНК может иметь место различное соединение экзонов друг с другом, что приведет к синтезу различных белков. Возможно, интроны служат материалом для образования новых генов в процессе эволюции. Показано, что мутации интронов могут нарушать процесс сплайсинга, останавливать синтез белка и изменять его структуру. Центральная догма молекулярной биологии: один ген – один фермент и ее современная трактовка. В сороковые годы Д. Бидл и Э. Татум выдвинули гипотезу: «Один ген – один фермент». Сегодня, несколько изменив первоначальную формулировку можно сказать: «Один ген – один белок или один ген – один полипептид». Хотя эта концепция сохраняет свое значение и по сей день, она уже недостаточно современная, т.к. известно, что существуют белки, которые кодируются генами, распределенными вдоль всего генома. Молекула ДНК (гена) выполняет различные функции. В ней имеются не только нуклеотидные последовательности, несущие генетическую информацию, но и такие, которые контролируют экспрессию (проявление) генов и репликацию. Механизмы генной регуляции у про- и эукариот. Экспрессия генов. Генетические механизмы экспрессии генов были изучены у микроорганизмов французскими генетиками Ф. Жакобом и Ж. Моно. Главное положение этой теории состоит в том, что в ДНК имеются следующие типы генов:
3) Транспозоны – это мобильные генетические элементы (мобильные ДНК, подвижные гены). Мобильные генетические элементы – это мобильные последовательности ДНК, найденные в геномах всех организмов. Во многих геномах они находятся в изобилии: например, они составляют до 50% человеческой ДНК. Большинство транспозонов способны встраиваться в различные участки ДНК. Они часто вызывают мутации, либо включаясь в другой ген и нарушая его нуклеотидную последовательность, или вызывая перестройки ДНК, такие как делеции, дупликации и инверсии. Мобильные элементы бывают автономными и неавтономными. Среди автономных, одни из них имеют только те последовательности, которые необходимы для их собственного перемещения, тогда как другие – имеют сложную структуру и кодируют ряд функций, не связанных непосредственно с перемещением. Неавтономные транспозоны для транспозиции нуждаются в ферментах, кодируемых автономными транспозонами. У человека транспозоны были обнаружены в 1991, когда Фрэнсис Коллинз и его коллеги обнаружили 31-летнего человека с нейрофиброматозом, вызванным перемещением последовательности Alu. Нейрофиброматоз – болезнь, которая вызывает многочисленные опухоли кожи и нервов. В настоящее время установлено, что от 45 до 50% (по данным разных авторов) человеческого генома состоят из последовательностей, происходящих от мобильных элементов, хотя большинство этих элементов является бездействующими и не способны к перемещению. Из них, около 2% – это ДНК транспозоны и приблизительно 42% – ретротраспозоны. Эволюционное значение мобильных генетических элементов неизвестно, но были предложены три гипотезы, объясняющие их происхождение. Гипотеза «клеточной функции» предполагает, что мобильные элементы обеспечивают какую-то важную функцию клетки. Гипотеза «генетической изменчивости» предполагает, что мобильные элементы, вызывая мутации, обеспечивают эволюционную гибкость видов. Гипотеза «эгоистичной ДНК» предполагает, что мобильные элементы не приносят какую-либо пользу клетке, но широко распространены из-за того, что они могут копироваться и распространяться. Один или несколько структурных генов, расположенных в бактериальной или вирусной «хромосоме» рядом с группой регуляторных генов, представляют вместе единицу генетической регуляции – оперон. Принципы работы оперона прокариот рассмотрим на примере работы оперона кишечной палочки (E. coli), ответственного за усвоение лактозы у этой бактерии (Рис 3).
Рис. 3 — Структура лактозного оперона Основу генетического аппарата кишечной палочки составляет бактериальная хромосома, входящая в состав нуклеоида. Нуклеоид этой бактерии включает различные участки, в том числе и лактозную область (lac оперон). Последняя область включает 3 гена, кодирующие 3 фермента: β галактозидазу, пермеазу и трансацетилазу (1, 2, 3), участвующих в метаболизме лактозы. Все гены laс оперона транскрибируются в одну и-РНК, которая транслируется с образованием 3-х белков. Оперон начинается с участка, к которому присоединяется особый белок-активатор – Сар-белок, активизирующий катаболические гены. Без этого белка фермент РНК-полимераза не может связаться с опероном и начать транскрипцию. Сар-белок предварительно активизируется сам присутствующим в клетке циклическим аденозинмонофосфатом (цАМФ). Вслед за этим участком лежит промотор. Это последовательность нуклеотидных пар, опознаваемая РНК-полимеразой. РНК-полимераза прикрепляется к промотору и затем продвигается вдоль оперона, транскрибируя его. За промотором находится оператор, состоящий из 21 пары нуклеотидов, который играет важную роль в регуляции работы оперона, так как с ним может связываться особый белковый фактор – регуляторный белок. Заканчивается laс оперон терминатором – небольшим участком ДНК, служащим стоп-сигналом, прекращающим продвижение РНК-полимеразы и транскрипцию оперона. Основная регуляция работы структурных генов laс оперона осуществляется регуляторным белком, который кодируется геном-регулятором. Этот белок синтезируется в клетке непрерывно, но в очень небольшом количестве (одновременно в цитоплазме присутствует не более 10 молекул). Регуляторный белок обладает сродством с оператором laс оперона, и если в питательной среде нет лактозы, то прикрепляется к оператору и препятствует продвижению РНК-полимеразы от промотора к структурным генам, которые оказываются репрессированными. Синтез кодируемых ферментов не идет. При поступлении в питательную среду лактозы регуляторный белок связывается с лактозой раньше, чем его молекулы достигнут оператора и сильно изменяет свою структуру, вследствие чего теряет способность присоединяться к оператору. РНК-полимераза свободно продвигается по оперону, транскрибирует структурные гены и в клетке начинается синтез всех трех ферментов, необходимых для усвоения лактозы, т.е. происходит индукция (экспрессия гена). При этом типе регуляции экспрессии генов лактоза выполняет роль эффектора – низкомолекулярного вещества, изменяющего свойства белка при соединении с ним. Регуляция активности генов у эукариот изучена менее полно, чем у вирусов и прокариот, что обусловлено наличием у них ядра, сложно устроенных хромосом и дифференциацией клеток. Допускается, что в основе регуляции действия генов у эукариот лежат механизмы, в принципе сходные с таковыми у вирусов и прокариот. Однако, есть и существенные отличия.
Примером сложной экспрессии генов может служить генный контроль синтеза гемоглобинов у человека. Известно, что гемоглобин человека является тетрамером, то есть состоит из четырёх субъединиц. У взрослого человека они представлены парными полипептидными цепями. Каждая цепь контролируется определенным генным локусом (Табл. 1). Таблица 1 — Генный контроль синтеза гемоглобинов у человека
НbА и НbА2 относятся к нормальным гемоглобинам человека. В эритроцитах плода около 80% гемоглобина представлено формой НbF, его молекула состоит из двух цепей α и двух цепей γ. У больных серповидноклеточной анемией имеется особый гемоглобин Нbs, который отличается от нормального НbА тем, что у него в одной β цепи в 6-ом положении глутаминовая кислота заменена валином. Существует мутантная форма гемоглобина – НbC. В нем в 6-ом положении глутаминовая кислота заменена лизином. Этот вариант гемоглобина назван «С» по названию города, у жителя которого была впервые обнаружена мутация – Christchurch (Новая Зеландия), хотя встречается преимущественно в Западной Африке. Четыре типа гемоглобинов контролируются отдельными генами: - локус αА определяет формирование α цепей в течение всей жизни у всех четырех гемоглобинов; - локус βА контролирует формирование β цепей только в НbА после рождения; - локус γF определяет синтез γ цепи в гемоглобине НbF в течение внутриутробной жизни; - локус σА2 определяет синтез σ цепей в гемоглобине НbА2 в течение всей жизни после рождения. Локусы αА, βА, σА2, γF тесно сцеплены в хромосоме. Все четыре указанных генов – структурные. В их действии имеется сложная экспрессия, благодаря чему возникают четыре типа гемоглобинов. Экспрессия генов βА, σА2 находится под влиянием генов-регуляторов. У взрослого человека происходит замена НbF плода на НbА или НbА2. При этом происходит репрессия гена γF и включение гена βА. Взаимодействие генов αА, βА, σА2 определяет развитие нормального гемоглобина и является примером межгенного взаимодействия. При формировании гемоглобина серповидноклеточной анемии наблюдается межаллельное взаимодействие аллели βА и ее патологической аллели. Вышеизложенные данные позволили сформулировать современную теорию гена, которая утверждает:
Генная инженерия. Одним из разделов молекулярной генетики и молекулярной биологии, который приблизился к рубежу практических приложений, является генная инженерия. Генная инженерия – это сумма методов, позволяющая переносить гены из одного организма в другой, или – это технология направленного конструирования новых биологических объектов (С.И. Алиханян, 1980 г.). К генной инженерии принято относить следующие операции:
Для того, чтобы организм обладал новыми наследственными свойствами необходимо ввести в него новый ген, подключить его к регуляторной системе организма, добиться его функционирования в соответствующих клетках. Существует 3 этапа такого процесса: получение генетического материала, введение генетического материала в изменяемый организм и включение новых генов в генетический аппарат клетки.
Осуществляется путем его выделения из генома клеток донора, либо путем его синтеза химическим путем или с помощью и-РНК и фермента обратной транскриптазы (ревертазы). Нужный ген получают путем вырезания его из ДНК с помощью особых ферментов – рестриктаз. Такой способ был разработан на плазмидной ДНК бактерий.
Осуществляется путем трансформации, трансдукции, конъюгации и соматической гибридизации. Гибридизация соматических клеток приводит к слиянию двух протопластов клеток и образованию ядра с диплоидным набором хромосом. Затем гибридная клетка делится, и образуются дочерние гибридные клетки с признаками обоих родителей.
Новые введенные гены не способны к самостоятельной репликации. Поэтому они вначале вводятся в состав структуры, способной к воспроизведению. Она называется вектором. Это молекула ДНК, которая способна переносить в клетку чужой ген и обеспечивать его размножение там. Векторами могут быть плазмиды бактерий, бактериофаги, вирусы животных, космиды – фрагменты бактериофагов с плазмидами. С помощью генной инженерии решено ряд проблем биологии: установлено мозаичное строение гена, расшифрована структура генов, кодирующих иммуноглобулины. В 1980 году получен гормон соматотропин, в 1982 – инсулин методом генной инженерии, а также интерферон, витамины, и различные гормоны. Генная инженерия является основой биотехнологии, а в медицине обеспечивает производство вакцин, сывороток, а также диагностику генетических аномалий человека на ранних стадиях развития (генная хирургия – замена поврежденного гена новым). В 2001 году усилиями генетиков программы «Геном человека» открыто количество генов, составляющих 31 тыс. на геном человека. Это расширяет возможности использования генетики для медицинских целей. Все работы по клонированию человека, по мнению большинства ученых-генетиков должны быть запрещены. Пересадка генов должна проводиться для человека только в терапевтических целях, с помощью соматических клеток. Применение половых клеток для генной терапии возможно только при достаточном доказательстве безопасности такого лечения по сравнению с генной терапией соматическими клетками. |