все лекции по биологии. Учебное пособие для студентов I курса, обучающихся по специальностям лечебное дело и медикодиагностическое дело
 Скачать 1.56 Mb. Скачать 1.56 Mb.
|

|
Тема № 4. Клеточный уровень организации живого. Клетка – элементарная генетическая и структурно-функциональная единица живого. Раздел биологии, занимающийся изучением структурной и функциональной организации клетки как единицы живого, получил название цитологии (от греч. cytos – клетка, полость, logos – наука). Открытие клетки связано с именами великих ученых-микроскопистов – Р. Гука, M. Мальпиги, Н. Грю, описавших ячеистое строение многих растительных объектов, а также с именем Антони Ван Левенгука, впервые наблюдавшего реальные клетки животных. В 1939 году немецкий зоолог Т. Шванн опубликовал труд "Микроскопические исследования о соответствии в структуре и росте животных и растений", в котором были заложены основы клеточной теории. В этой работе Т. Шванн пришел к двум выводам: 1) клетка – главная структурная единица всех растительных и животных организмов; 2) процесс образования клеток обусловливает рост, развитие и дифференцировку всех растительных и животных тканей и организмов. Дальнейшее развитие клеточной теории связано с именем немецкого ученого Рудольфа Вирхова, который в 1858 году опубликовал свой труд «Целлюлярная патология». В этой работе Р. Вирхов дополнил клеточную теорию третьим выводом: «Omnis cellula e cellula» каждая клетка из клетки. Этот вывод блестяще подтвердился дальнейшим развитием биологии. В настоящее время не известно иных способов появления клеток помимо их деления. В своей работе Р. Вирхов впервые подошел к объяснению патологического процесса, показав его связь в организме с морфологическими структурами, с определенными изменениями в структуре и функции клеток. Он является основоположником патологической анатомии. Однако, ряд выводов Р. Вирхова оказались ошибочными и закономерно встретили возражения со стороны современников. По Р. Вирхову патологический процесс в организме представляет собой сумму нарушений жизнедеятельности отдельных клеток, это локальный процесс. Р. Вирхов и его последователи не видели также качественных отличий между частью и целым, рассматривая организм вне его исторического развития и условий существования. Эту идею Р. Вирхова обоснованно критиковали И.М. Сеченов, C.П. Боткин, И.П. Павлов, которые показали, что организм – единое целое и интеграция его частей осуществляется прежде всего ЦНС. Благодаря исследованиям Т. Шванна, М. Шлейдена, Р. Вирхова, Т. Моргана, С.Г. Навашина, Н.К. Кольцова, Д.Н. Насонова и др., клетку рассматривают как наименьшую элементарную единицу живого, которой свойственны такие признаки, как метаболизм, воспроизведение, реактивность и изменчивость. Клеточная теория, основные этапы ее развития. Современное состояние клеточной теории. Современная клеточная теория включает следующие положения: 1. Клетка – основная структурно-функциональная и генетическая единица живого. 2. Клетки одно- и многоклеточных организмов сходны по строению, химическому составу и проявлению жизнедеятельности. 3. Размножение клеток осуществляется путем деления исходной (материнской) клетки. 4. Клетки многоклеточных организмов специализируются по функциям и образуют ткани и органы. 5. Единое целое организма и интеграция его частей осуществляется, прежде всего, ЦНС. 6. В основе непрерывности, единства и разнообразия органического мира лежат обмен веществ, размножение, наследственность, изменчивость и раздражимость клеток. Значение клеточной теории: - доказательство морфологической основы единства живой природы; - общебиологическое объяснение живой природы; - доказательство эволюционных процессов. Доклеточные формы живого. Жизненные формы организмов:
эукариоты – царства растений, животных и грибов. Большинство живых организмов состоит из клеток. Однако имеются неклеточные формы жизни – вирусы. С возникновением клетки живые системы приобретают способность к самостоятельному обмену веществ и размножению. Усложнение их организации связано с появлением клеточной, а затем ядерной мембраны и увеличением молекулярной массы ДНК. Особенности строения прокариотической клетки. Прокариоты – одноклеточные доядерные организмы. Наследственный аппарат представлен одной молекулой ДНК кольцевой формы. ДНК вместе с белками формирует в бактериальной клетке особый комплекс – нуклеоид. Прокариоты являются гаплоидами. Молекулярная масса ДНК соответствует приблизительно 2000 структурных генов. Клетка ограничена двойной плазматической мембраной (наружной и внутренней). Поверх мембраны образуется клеточная стенка. Она состоит из углевода муреина, образующего жесткую решетку. В цитоплазме отсутствуют органеллы мембранного строения. Их функцию выполняют впячивания внутренней мембраны – мезосомы. В цитоплазме имеются рибосомы. Бактерии могут содержаться мелкие молекулы ДНК (плазмиды). Фотосинтезирующие бактерии имеют фотомембраны. Запасные питательные вещества представлены углеводами. Структурные компоненты клеток эукариот: плазматическая мембрана, цитоплазма, ядро. Их строение. Классификация органоидов и включений. Эукариотические клетки имеют обособленное ядро, наружную биологическую мембрану – плазмолемму и цитоплазму с органеллами и включениями. Плазмолемма отделяет содержимое клетки от внешней среды и регулирует движение ионов и макромолекул в клетку и из нее. Плазмолемма имеет жидкостно-мозаичное строение (модель Сингера). Она состоит из двойного фосфолипидного слоя, белков и полисахаридов. Молекулы фосфолипидов представлены неполярными гидрофобными концами и полярными гидрофильными головками, обращенными к внешней среде. Белки расположены мозаично: поверхностные, погруженные и пронизывающие. На поверхности мембраны находятся олигосахаридные цепи, состоящие из моносахаридных остатков. Их функции:
Клетки растений имеют целлюлозную, а грибов – хитиновую оболочки поверх плазмолеммы. На наружной поверхности плазмолеммы животных клеток находится полисахаридный слой – гликокаликс. Химический состав клеточной мембраны следующий: 1) белки – 55% (из них до 200 ферментов); 2) липиды – 35%; 3) углеводы – 5-10% (в соединении с простыми или сложными белками). Функции липидов мембран: структурная, барьерная. Функции белков мембран: структурная, ферментативная, рецепторная, транспортная. Функция гликопротеидов – рецепторная. Свойства мембран: пластичность, полупроницаемость, динамичность. Функции мембран: 1) структурная (входят в состав большинства органоидов); 2) барьерная (поддерживает постоянство химического состава) и защитная; 3) регуляторная (регуляция обменных процессов); 4) рецепторная; 5) транспортная. Через плазмолемму осуществляется транспорт веществ в клетку. Транспорт бывает пассивный и активный. 1. Пассивный транспорт происходит без затрат энергии, по градиенту концентрации. Это может быть: диффузия газов, осмотическое движение воды, облегченная диффузия веществ (аминокислот, сахаров, жирных кислот) посредством белков-переносчиков. 2. Активный транспорт идет против градиента концентрации, с затратой энергии. Для него необходимо наличие специальных ионных каналов, ферментов и АТФ. Так работает натрий-калиевый насос. Концентрация калия в клетке выше, чем в околоклеточном пространстве, и, тем не менее, ионы калия поступают в клетку, а ионы натрия выводятся наружу. Ионы натрия формируют на поверхности мембраны положительный заряд, внутри клетки заряд отрицательный по отношению к среде. На каждые 2 поступающие иона К+ из клетки выводится 3 иона Na+. Заряд на мембране обеспечивает передачу нервного импульса, всасывание питательных веществ ворсинками кишечника, адсорбцию в почечных канальцах. Mg2+/Ca2+ насос обеспечивает мышечные сокращения. Крупномолекулярные соединения белков, нуклеиновых кислот, полисахаридов проникают внутрь клетки путем эндоцитоза. Различают два вида эндоцитоза: фагоцитоз и пиноцитоз. Фагоцитоз – захватывание мембранной твердых частиц. При этом внутри клетки при участии лизосом образуется пищеварительная вакуоль. Пиноцитоз – поступление жидкостей в клетку. Выделение из клетки веществ, заключенных в мембрану, называется экзоцитозом. Поступившие в клетку вещества могут использоваться: 1) для синтеза веществ, необходимых самой клетке (анаболическая система); 2) как источник энергии (катаболическая система). Цитоплазма – живое содержимое клетки без ядра. В цитоплазме различают гиалоплазму, органеллы и включения. Гиалоплазма является основным веществом клетки, с которым связаны коллоидные свойства цитоплазмы, ее вязкость, эластичность, сократимость и внутреннее движение. Гиалоплазма состоит из двух фаз: жидкой и твердой. Жидкая фаза представлена коллоидным раствором белков, углеводов, нуклеотидов и ионов неорганических веществ. Твердая фаза представлена микротрабекулярной системой, микротрубочками и микрофиламентами (фибриллы), которые образуют цитоскелет клетки. Органеллы – это специализированные постоянные компоненты цитоплазмы, обладающие определенным строением и выполняющие ту или иную функцию в жизнедеятельности клетки. Органеллы подразделяют на две группы: общего и специального назначения. 1. Органеллы общего назначения (встречаются во всех видах клеток): а) мембранного строения (митохондрии, пластиды, комплекс Гольджи, эндоплазматический ретикулум, лизосомы, вакуоли); б) немембранного строения (рибосомы, клеточный центр). 2. Органеллы специального назначения (характерны для определенного типа клеток): миофибриллы, тонофибриллы, нейрофибриллы, реснички, жгутики и микроворсинки. Цитоплазматические включения – это непостоянные структуры в цитоплазме, представляющие собой продукты жизнедеятельности клеток. По своему биологическому значению включения могут быть условно разделены на основные группы: трофические, секреторные, специального назначения, экскреторные, пигментные. Ядро (nucleus, karion) –это постоянный структурный компонент всех клеток эукариот. Оболочка интерфазного ядра состоит из двух элементарных мембран (наружной и внутренней), пространство между которыми называется перинуклеарным. Мембраны имеют поры, через которые идет обмен веществ между ядром и цитоплазмой. Наружная ядерная мембрана переходит в стенки каналов гранулярной эндоплазматической сети, на которой расположены рибосомы. Нуклеоплазма –однородная масса, заполняющая пространство между структурами ядра. Она содержит белки, нуклеотиды, АТФ и различные виды РНК, а так же хроматин и ядрышки. Хроматин – комплекс ДНК и гистоновых белков (дезоксирибонуклеопротеид) в отношении 1: 1,3. Выявляется в интерфазном ядре. В процессе митоза спирализованный хроматин образует хромосомы. Ядрышко – состоит из белков (80%), РНК (до 15%), ДНК (до 12%). Образуется в ядрышковом организаторе на хромосоме. Обеспечивает синтез р-РНК и образование субъединиц рибосом. Функции ядра: хранение и передача генетической информации, регуляция процессов жизнедеятельности клетки. Анаболическая система клетки и ее органоиды: эндоплазматический ретукулюм, комплекс Гольджи, рибосомы. Анаболическая (ассимиляция, пластический обмен) и катаболическая (диссимиляция, энергетический обмен) системы клетки неразрывно связаны, так как все процессы жизнедеятельности клетки немыслимы без энергии АТФ, которая, в свою очередь, не может образовываться без ферментных систем, строящихся в результате анаболических реакций. Также неразрывно связаны друг с другом потоки вещества и энергии, так как гетеротрофные клетки способны использовать только энергию, заключенную в сложных химических соединениях. К анаболической системе клетки относятся: рибосомы, эндоплазматический ретикулум, комплекс Гольджи. Катаболическая система и ее органоиды: лизосомы, пероксисомы, глиоксисомы, митохондрии. К катаболической системе клетки относятся: лизосомы, пероксисомы, глиоксисомы, митохондрии. Пероксисомы – клеточные органеллы, в которых осуществляются окисление жирных кислот, синтез желчных кислот, холестерина, а так же эфиросодержащих липидов, участвующих в построении миелиновой оболочки нервных волокон. Они есть во всех эукариотических клетках. Их функции сильно различаются в клетках разных типов. Это один из главных центров утилизации кислорода в клетке. Содержат ферменты: оксидазы, уратоксидазы и каталазы. Каталаза окисляет фенолы, муравьиную кислоту, формальдегид и спирты. Этот тип окислительных реакций особенно важен в клетках печени и почек, где пероксисомы обезвреживают ядовитые вещества, попадающие в кровоток. Глиоксисомы – клеточные органеллы растений, которые содержат ферменты, необходимые для превращения жиров в углеводы. Они принимают метаболиты, поступающие из жировых капель – сферосом, превращают их в янтарную кислоту, которая затем подвергается последовательному превращению в продукты, восстанавливаемые до сахаров. Тема № 5. Обменные процессы в жизненном цикле клетки. Клетка – открытая система. Клетка является открытой, саморегулирующейся системой. Для нее характерен поток вещества, энергии и информации, обеспечивающие обменные процессы. Для клетки, как и в целом для организма, различают внешний и внутренний обмен. Внешний обмен – это обмен с внешней средой: поступление питательных веществ, выделение продуктов метаболизма. Внутренний обмен осуществляется путем катаболизма (диссимиляции) и анаболизма (ассимиляции). Ассимиляция осуществляется путем реакций пластического обмена (биосинтез белков, жиров, углеводов, фотосинтез). По типу ассимиляции организмы делят на автотрофные и гетеротрофные. Диссимиляция осуществляется путем реакций энергетического обмена (синтез АТФ за счет энергии расщепления сложных органических веществ). По типу диссимиляции организмы бывают анаэробные и аэробные. Организации энергетического обмена в клетке. Энергия – это «способность производить внешнее действие» (М. Планк, т.е. совершать работу). По виду обмена веществом или энергией с окружающей средой различают следующие виды систем: изолированные (невозможен обмен веществ и энергии), адиабатические (невозможен обмен веществ, но возможен обмен энергией кроме тепловой), замкнутые (невозможен обмен веществ, но возможен обмен энергией в любой форме) и открытые (возможен любой обмен веществом и энергией) системы. Все клетки и живые организмы являются гетерогенными открытыми системами. Первичным источником энергии для жизни на Земле является солнечная энергия. Для всех клеток живых организмов органические вещества (углеводы, жиры и частично белки) с их химической энергией служат источником энергии, необходимой для жизнедеятельности организма. Автотрофные организмы сами синтезируют энергию в таком виде, гетеротрофные – получают ее от автотрофных. Энергия химических связей, освобождающаяся при распаде органических веществ, не используется в клетках сразу для осуществления работы, а превращается в энергию макроэргических связей молекулы АТФ. В процессе фотосинтеза солнечная энергия в растительных клетках сначала превращается в энергию молекул АТФ, НАДФН+Н+, а затем молекул органических веществ. В клетках гетеротрофных организмов поток энергии начинается с поступления готовых органических веществ, а затем обеспечивается процессами брожения и дыхания. Брожение происходит в гиалоплазме в анаэробных условиях. При этом промежуточными продуктами диссимиляции органических веществ являются две молекулы пировиноградной кислоты (С3Н4О3) и две молекулы АТФ. Поэтому, выход энергии при брожении невелик. Он составляет 40%, а 60% – рассеивается в виде тепла. Высокоэнергетичным процессом является дыхание, при котором в матриксе и на мембранах крист митохондрий образуется еще 36 молекул АТФ. В сумме энергия одной молекулы глюкозы трансформируется в энергию 38 молекул АТФ. Энергетический выход на кислородном этапе – 36 АТФ (цикл Кребса – 2 АТФ, электронтранспортная цепь 34 АТФ) (Рис. 4). Процесс энергетического обмена включает: подготовительный этап, бескислородный и кислородный. На первом этапе наблюдается образование мономеров из сложных органических веществ. Во втором – участвует более 10 ферментативных реакций цитоплазмы. На этом этапе завершается энергетический обмен организмов анаэробов. При этом, из АДФ и фосфатов клетки синтезируется только 2 молекулы АТФ и 2 молекулы пировиноградной кислоты. Световая фаза Темновая фаза 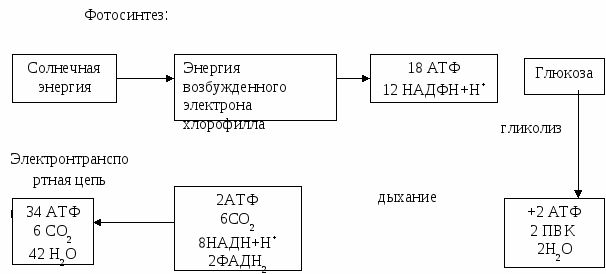 hυ  Цикл Кребса  Рис. 4 — Общая схема потока энергии У высокоорганизованных животных и человека гликолиз является обязательным дополнительным источником энергии к аэробиозу. При недостатке кислорода в мышцах продолжается анаэробное расщепление пировиноградной кислоты. При этом пировиноградная кислота превращается в молочную. Полное извлечение энергии из промежуточного продукта распада пировиноградной кислоты происходит на кислородном этапе, включающем цикл Кребса в матриксе митохондрий и дыхательную цепь на мембранах крист (10 реакций последовательного превращения трикарбоновых кислот). В цикле Кребса в результате последовательного превращения образуется 2 молекулы АТФ, 6СО2, 8НАДН+Н+ и 2ФАДН2. Последние являются переносчиками водорода к мембранам крист. В дыхательной цепи образуются 34 АТФ и конечный продукт распада – вода. Всего на кислородном этапе энергетический выход составляет 36 АТФ. Общий выход энергии – 38 молекул АТФ из одной молекулы глюкозы. На синтез АТФ клетка использует 67% энергии поступающих в нее органических веществ. Пластический обмен в клетке в процессе фотосинтеза, хемосинтеза и биосинтеза белка. Структуры живых организмов непостоянны. Они разрушаются и образуются заново. Процесс обновления осуществляется за счет потока вещества. Поток вещества характеризуется пластическим обменом в клетке: 1. Фотосинтез – это синтез органического вещества из неорганических в хлоропластах с использованием солнечной энергии. h 6СО2 + 6Н2О → С6Н12О6 + 6О2 2. Хемосинтез. При хемосинтезе происходит преобразование энергии окисления неорганических веществ в энергию химических связей органических веществ. Энергия, которая выделяется при окислении соединений: сероводорода (серные бактерии), аммиака (нитрифицирующие бактерии), закиси железа (железобактерии), используется для восстановления двуокиси углерода до органических соединений. 3. Биосинтез белка. Процесс переноса информации о структуре белка с и-РНК на синтезируемый полипептид называется трансляцией. Трансляция происходит в цитоплазме на рибосоме и включает следующие процессы: - активация аминокислоты специфическим ферментом – аминоацил т РНК-синтетазой, в присутствии АТФ с образованием аминоациладенилата (тройной комплекс); - присоединение активированной аминокислоты к специфической т РНК с высвобождением аденозинмонофосфата (АМФ); - связывание этого комплекса с рибосомами; - включение аминокислот в белок с высвобождением т-РНК. Рибосома контактирует в каждый момент только с небольшим участком и-РНК, возможно равным одному триплету (кодону). Рибосома движется по и-РНК кодон за кодоном. На и-РНК одновременно может быть несколько рибосом, образующих полисому. В рибосому, движущуюся по и-РНК, т-РНК поставляет активированные аминокислоты. Аминокислота прикреплена к т-РНК, на изгибе средней петли последней имеется триплет, получивший название антикодона. При совпадении антикодона т-РНК с кодоном и-РНК, по принципу комплементарности, аминокислота включается в полипептидную цепь. Поток информации в клетке. Каждой клетке, живому организму свойственен обмен информации. На уровне клетки организм получает информацию о том, что происходит в окружающей среде. Непрерывный поток информации поступает в организм и там перерабатывается, другой такой поток информации все время выходит из организма, служащего, в свою очередь, передатчиком. При гормональной связи химические сигналы в виде гормонов попадают в организм, но только определенные клетки – мишени специфических органов способны принять данный сигнал. Так, тиреотропный гормон гипофиза регулирует функцию щитовидной железы. В нервной системе информационным параметром служит число импульсов в единицу времени (частота импульсов). Количество информации в том или ином сигнале измеряют в битах. У человека максимальный поток информации, направленный, главным образом, по зрительному анализатору внутрь, оценивается в 108 – 109 бит/с, что составляет 109 бит за 80 лет жизни (для сравнения: книга среднего объема содержит около 106 бит). Для управления поведением человека, активностью его физиологических функций необходим выходной (направление из мозга) поток информации объемом 107 бит/с. Он обеспечивается подключением программ памяти. Поток внутренней информации в организме осуществляется с помощью коры головного мозга и желез внутренней секреции. Клетка воспринимает поток внешней информации из межклеточной жидкости при помощи рецепторов клеточной мембраны. Внутренняя информация содержится в ДНК. В потоке внутренней информации принимает участие ядерная и цитоплазматическая ДНК, РНК, ферменты. Поток информации обеспечивает наследственную преемственность поколений. Жизненный цикл клетки. Авторепродукция клеток. Период времени от деления материнской клетки до собственного деления дочерней или ее гибели называют жизненным циклом (или клеточным). Для клеток неделящихся клеточных популяций клеточный цикл составляет период от образования клетки до ее смерти (тела нейронов, клетки миокарда). В жизненном цикле клеток размножающейся клеточной популяции можно различить два больших периода: 1. Интерфаза – период между делениями, когда клетка растет, функционирует, подготавливается к делению; 2. Митоз – деление клетки. Интерфаза имеет три периода: 1. Постмитотический или пресинтетический период G1. В этот период клетки растут, синтезируют РНК, белки, накапливается энергия, но синтез ДНК отсутствует. Период характеризуется тем, что клетка содержит диплоидный набор хромосом (2n). Каждая хромосома образована одной хроматидой. Хромосомы деспирализованы. Если количество ДНК, содержащееся в гаплоидном наборе хромосом человека обозначить "С", то в G1 количественная характеристика генетического материала: 2n 1 хроматида 2С. 2. Синтетический период S: В клетках идет активный синтез ДНК, хромосомных структур, РНК и беков-гистонов. Каждая хромосома достраивает вторую хроматиду. Количественная характеристика генетического материала: 2n 2 хроматиды 4С. 3. Постсинтетический, или премитотический период G2: В клетке идет синтез белков митотического аппарата, накапливается энергия. Генетическая характеристика: 2n 2 хроматиды 4С. Одной из основных биологических особенностей клетки как элементарной живой системы является ее способность к авторепродукции. Репродукция клеток лежит в основе развития, роста, размножения и регенерации организма. Митоз – наиболее распространенный способ репродукции клеток (табл. 2). Универсальность этого способа деления клеток связана с тем, что он обеспечивает образование генетически равноценных клеток и сохраняет преемственность хромосом в ряду клеточных поколений. Таблица 2 — Типы и виды деления клеток
Биологическое значение митоза:
В процессе митоза идет последовательная смена фаз: профаза, прометафаза, метафаза, анафаза, телофаза. Профаза – происходит спирализация, укорочение, утолщение хроматиновых нитей. Наблюдается расхождение центриолей к полюсам. Начинают образовываться нити веретена деления. В конце – наблюдается разрушение ядрышка и ядерной оболочки. Количественная характеристика генетического материала: 2n 2 хроматиды 4С. Метафаза – хромосомы в животных клетках располагаются в упорядоченном состоянии в области экватора. Образуется метафазная пластинка. Завершается образование нитей веретена деления. Нити веретена, которые крепятся к кинетохорам, называются хромосомными, а которые идут от полюса к полюсу – полюсными. Количественная характеристика генетического материала: 2n 2 хроматиды 4С. Анафаза – хромосомные нити веретена сокращаются. К противоположным полюсам расходятся хроматиды, которые принято называть дочерними хромосомами. На каждом полюсе количественная характеристика генетического материала: 2n 1хроматида 2С. Телофаза – дочерние хромосомы, разошедшиеся к полюсам, деспирализуются, теряют ясные очертания, вокруг них формируются ядерные оболочки, восстанавливается ядрышко. Клеточный центр теряет активность. Начинается цитокинез – деление цитоплазмы. Итогом деления является образование двух диплоидных клеток. Деление в растительной и животной клетках происходит сходно. Но в клетках высших растений отсутствует клеточный центр. Цитотомия в животных клетках происходит путем перетяжки (образование борозды), которая, углубляясь, делит клетку на две части. В клетках растений формируется в центре срединная пластинка, которая затем растет к периферии. Митотический цикл клетки – совокупность процессов подготовки клеток к делению и само митотическое деление. Если дочерние клетки, или клетка, сразу же приступают к подготовке к следующему митозу, то их митотический цикл совпадает с жизненным циклом (ткани эмбриона). В других случаях дочерние клетки подвергаются дифференцировке и выполняют различные функции (пресинтетический период удлиняется). Их жизненный цикл заканчивается смертью клетки (у нервных клеток G1 – в течение всей жизни). Продолжительность каждого из периодов митотического цикла и фаз митоза различна и длится от нескольких минут до нескольких часов, что зависит от ряда причин: типа тканей, физиологического состояния организма, внешних факторов (t, свет, химические вещества). Оказывают влияние и факторы внутренней среды: нейрогуморальные механизмы, осуществляемые нервной системой и гормонами, а также продукты распада тканей. Важную роль играют факторы, обеспечивающие возможность вступления клеток в деление. Четко доказано, что все синтетические процессы в клетке, готовящейся к делению, находятся под контролем ее генетического аппарата. Гены, контролирующие этот процесс, находятся в разных хромосомах. Активность генов объясняется гипотезой Ф. Жакоба и Ж. Моно (1961). На важную роль нейрогуморальной регуляции митотической активности указали ученые Л. Н. Бляхер (1954), И. А. Уткин (1959). Они установили, что рефлекторный характер регуляции клеточных делений влияет опосредованно – через сдвиг гормонального равновесия. Установлено, что усиление секреции адреналина тормозит митотическую активность, тогда как гормоны щитовидной железы вызывают усиление митоза. Удаление надпочечников приводит к выключению эффекта торможения митоза. На митотический цикл также влияют: суточный ритм митотический активности, факторы внешней среды (свет, температура) нейрогуморальные механизмы, продукты распада тканей. Циклины − семейство белков, являющихся активаторами циклин-зависимых киназ (CDK) (CDK — англ. cyclin-dependent kinases) — ключевых ферментов, участвующих в регуляции клеточного цикла эукариот. Циклины получили свое название в связи с тем, что их внутриклеточная концентрация периодически изменяется по мере прохождения клеток через клеточный цикл, достигая максимума на его определенных стадиях. Каталитическая субъединица CDK частично активируется в результате взаимодействия с молекулой циклина, которая образует регуляторную субъединицу фермента. Образование этого гетеродимера становится возможным после достижения циклином критической концентрации. В ответ на уменьшение концентрации циклина происходит инактивация фермента. Для полной активации CDK должно произойти специфическое фосфорилирование и дефосфорилирование определенных аминокислотных остатков в полипептидных цепях этого комплекса. Одним из ферментов, осуществляющих подобные реакции, является киназа CAK (CAK — CDK активирующая киназа). Циклин-зависимые киназы – группа белков, регулируемых циклином и циклиноподобными молекулами. Большинство CDK участвуют в смене фаз клеточного цикла; также они регулируют транскрипцию и процессинг мРНК. Известно несколько CDK, каждая из которых активируется одним или более циклинами и иными подобными молекулами, притом по большей части CDK гомологичны, отличаясь в первую очередь конфигурацией сайта связывания циклинов. Эндомитоз – один из видов митоза, суть которого заключается в редупликации хромосом без разрушения ядерной оболочки и без деления клетки (образование полиплоидов). Вследствие этого в клетке происходит умножение числа хромосом, иногда в десятки раз по сравнению с исходным. Эндомитоз встречается в интенсивно функционирующих клетках различных тканей: тканях нематод, насекомых, ракообразных, в корешках некоторых растений. Допускают, что эндомитоз возникает в процессе эволюции, как один из вариантов митоза. Политения – многократное воспроизведение в хромосомах количества хромонем без увеличения числа хромосом в клетке. При политении выпадают все фазы митотического цикла, кроме репродукции хромонем. Политения встречается у двукрылых насекомых, инфузорий и у некоторых растений. Используется для построения карт хромосом, а также обнаружения хромосомных перестроек. Мейоз – деление, обеспечивающее образование гаплоидных клеток из соматических. Значение мейоза: 1. Обеспечивает образование половых клеток с гаплоидным набором хромосом. 2. Поддерживает постоянное число хромосом в кариотипе. 3. Обуславливает образование большого количества новых комбинаций генов, что является источником комбинативной изменчивости. 4. Обеспечивает возможность полового размножения. Состоит из двух последовательных делений: 1. Мейоз I – редукционное; 2. Мейоз II – эквационное. Мейоз I. Профаза 1 имеет 5 стадий: лептотена, зиготена, пахитена, диплотена и диакинез. Количественная характеристика генетического материала: 2n 2хроматиды 4С. Лептотена – хромосомы приобретают форму нитей, различимых в микроскоп. Зиготена – конъюгация гомологичных хромосом, образование бивалентов. Пахитена – происходит обмен участками гомологичных хромосом – кроссинговер. Диплотена – отталкивание между гомологичными хромосомами в области центромер. Хромосомы остаются связанными в области перекреста. Эти места называются хиазмами. Диакинез – спирализация максимальная, биваленты располагаются по периферии ядра. Разрушаются ядрышко и ядерная оболочка. Центриоли расходятся к полюсам и начинает образовываться веретено деления. Метафаза 1 – биваленты выстраиваются в экваториальной плоскости, центромерами прикрепляются к нитям веретена деления. Количественная характеристика генетического материала: 2n 2хроматиды 4С. Анафаза 1 – расхождение гомологичных хромосом к полюсам клетки. На каждом полюсе формируется гаплоидный набор хромосом. Каждая хромосома состоит из 2 хроматид. Количественная характеристика генетического материала: n 2хр. 2С. Телофаза 1 – характерна для клеток животных при этом образуются 2 клетки с гаплоидным набором. Клетки растений сразу переходят в мейоз II. Между мейозом I и мейозом II наблюдается интеркинез, в котором репликация ДНК отсутствует. Мейоз II –точная копия митоза. Профаза 2 – непродолжительная. Метафаза 2 – образование экваториальной пластинки. Анафаза 2 – расхождение сестринских хроматид. Количественная характеристика генетического материала: n 1 хроматида 1С. Телофаза 2 – формирование ядер, деление цитоплазмы и образование 4 гаплоидных клеток. Количественная характеристика генетического материала: n 1 хроматида 1С. Амитоз, или прямое деление, представляет собой деление ядра без формирования аппарата деления и спирализации хромосом. Хромосомы распределяются произвольно между дочерними клетками. Прямое деление характеризуется первоначально перешнуровкой ядрышка, затем ядра и цитоплазмы. Ядро может делиться на две равномерные части – равномерный амитоз, или две неравномерные части – неравномерный амитоз, либо ядро делится на несколько частей – фрагментация, шизогония. Иногда после деления ядра цитоплазма не делится, и возникают многоядерные клетки – амитоз без цитотомии. В зависимости от факторов, обуславливающих амитоз, выделяют три его вида: генеративный, реактивный, дегенеративный. Генеративный амитоз отмечается при делении высокоспециализированных полиплоидных клеток. Наблюдается у инфузории при делении макронуклеуса, а также в некоторых клетках млекопитающих (печени, эпидермиса). Реактивный амитоз выявляется при различных повреждающих воздействиях: ионизирующего облучения, нарушении обменных процессов, голодании, нарушении нуклеинового обмена и денервации ткани. Этот вид амитоза обычно не завершается цитотомией и приводит к образованию многоядерных клеток. Вероятно, его следует рассматривать как компенсаторную реакцию, приводящую к увеличению поверхности обмена между ядром и цитоплазмой. Дегенеративный амитоз возникает в стареющих клетках с угасающими жизненными свойствами. Этот вид представлен фрагментацией и почкованием ядер. Он не имеет отношения к репродукции клеток. Появление дегенеративных форм амитоза служит одним из признаков некробиотических процессов. Прямое бинарное деление характерно для прокариот. Включает репликацию кольцевой ДНК и далее – деление цитоплазмы с образованием двух клеток. Клеточная пролиферация и ее значение для медицины. Клеточная пролиферация – увеличение числа клеток путем митоза, приводящее к росту ткани, в отличие от другого способа увеличения ее массы (например, отек). У нервных клеток пролиферация отсутствует. Во взрослом организме продолжаются процессы развития, связанные с делением и специализацией клеток. Эти процессы могут быть как нормальными физиологическими, так и направленными на восстановление организма вследствие нарушения его целостности. Значение пролиферации в медицине определяется способностью клеток разных тканей к делению. С делением клеток связан процесс заживления ран и восстановление тканей после хирургических операций. Пролиферация клеток лежит в основе регенерации (восстановления) утраченных частей. Проблема регенерации представляет интерес для медицины, для восстановительной хирургии. Различают физиологическую, репаративную и патологическую регенерацию. Физиологическая – естественное восстановление клеток и тканей в онтогенезе. Например, смена эритроцитов, клеток кожного эпителия. Репаративная – восстановление после повреждения или гибели клеток и тканей. Патологическая – разрастание тканей не идентичных здоровым тканям. Например, разрастание рубцовой ткани на месте ожога, хряща – на месте перелома, размножение клеток соединительной ткани на месте мышечной ткани сердца, раковая опухоль. В последнее время принято разделять клетки тканей животных по способности к делению на три группы: лабильные, стабильные и статические. К лабильнымотносятся клетки, которые быстро и легко обновляются в процессе жизнедеятельности организма (клетки крови, эпителия, слизистой ЖКТ, эпидермиса и др.). К стабильным относятся клетки таких органов как печень, поджелудочная железа, слюнные железы и др., которые обнаруживают ограниченную способность к делению. К статическим относятся клетки миокарда и нервной ткани, которые, как считает большинство исследователей, не делятся. Изучение физиологии клетки имеет важное значение для понимания онтогенетического уровня организации живого и механизмов саморегуляции клетки, обеспечивающих целостное функционирование всего организма. | |||||||||||||||||||||