Генетика. Успехи современной генетики
 Скачать 0.77 Mb. Скачать 0.77 Mb.
|

|
Введение Основой современной биологии является Генетика, т.к. она изучает два неразрывных свойства живых организмов: наследственность и изменчивость. Однако роль генетики не ограничивается сферой биологии. Поведение человека, экология, социология, психология, медицина - вот далеко не полный список научных направлений, прогресс которых зависит от уровня знаний в области генетики. С учетом «сферы влияния» генетики понятна ее методологическая роль. В настоящее время именно генетика определяет единство биологических наук, благодаря универсальности законов наследственности и фундаментальной информации, систематизированной в положениях общей генетики. Методологическая роль генетики в полной мере распространяется на все науки о человеке. Живая природа устроила себя гениально, просто и мудро. У нее есть единственная самовоспроизводящая молекула ДНК, на которой записана программа жизни, а конкретнее, весь процесс синтеза, структура и функция белков, как основных элементов жизни. Кроме сохранения программы жизни, молекула ДНК выполняет еще одну важнейшую функцию - ее самовоспроизведение, копирование, создает преемственность между поколениями, непрерываемость нити жизни. Единожды возникнув, жизнь самовоспроизводится в огромном разнообразии, которое обеспечивает ее устойчивость, приспособленность к разнообразным условиям среды и эволюцию. Успехи современной генетики Современная генетика обеспечила новые возможности для исследования деятельности организма: с помощью индуцированных мутаций можно выключать и включать почти любые физиологические процессы, прерывать биосинтез белков в клетке, изменять морфогенез, останавливать развитие на определенной стадии. Мы теперь можем глубже исследовать популяционные и эволюционные процессы, изучать наследственные болезни, проблему раковых заболеваний и многое другое. В последние годы бурное развитие молекулярно-биологических подходов и методов позволило генетикам не только расшифровать геномы многих организмов, но и конструировать живые существа с заданными свойствами. Таким образом, генетика открывает пути моделирования биологических процессов и способствует тому, что биология после длительного периода дробления на отдельные дисциплины вступает в эпоху объединения и синтеза знаний Международные проект «Геном человека» был начат в 1988 г. Это один из самых трудоемких и дорогостоящих проектов в истории науки. Основная цель проекта было изучение последовательности нуклеотидных оснований во всех молекулах ДНК человека и установление локализации, т.е. полное картирование всех генов человека. Что же представляет собой основной предмет проекта - геном человека? Известно, что в ядре каждой соматической клетки (кроме ядра ДНК есть еще и в митохондриях) человека содержится 23 пары хромосом, каждая хромосома представлена одной молекулой ДНК. Суммарная длина всех 46 молекул ДНК в одной клетке равна приблизительно 2 м, они содержат около 3,2 млрд пар нуклеотидов. Общая длина ДНК во всех клетках человеческого тела (их примерно 5х1013) составляет 1011 км, что почти в тысячу раз больше расстояния от Земли до Солнца. Как же помещаются в ядре такие длиннющие молекулы? Оказывается, в ядре существует механизм «насильственной» укладки ДНК в виде хроматина -уровни компактизации. В ходе выполнения проекта «Геном человека» было разработано много новых методов исследования, большинство из которых в последнее время автоматизировано, что значительно ускоряет и удешевляет работу по расшифровке ДНК. Эти же методы анализа могут использоваться и для других целей: в медицине, фармакологии, криминалистике и т.д. В мире каждый сотый ребенок рождается с каким-либо наследственным дефектом. К настоящему времени известно около 10 тыс. различных заболеваний человека, из которых более 3 тыс. - наследственные. Уже выявлены мутации, отвечающие за такие заболевания, как гипертония, диабет, некоторые виды слепоты и глухоты, злокачественные опухоли. Обнаружены гены, ответственные за одну из форм эпилепсии, гигантизм и др. В последние годы была открыта молекулярная основа наследственности, расшифрован генетический код; создаются новые искусственные гены; в пробирках выращиваются вирусы; из клеток зрелого организма создаются идентичные близнецы лягушек и овец; в пробирках оплодотворяются человеческие клетки; женщинам пересаживают эмбрионы; врачи лечат многие наследственные заболевания; выращиваются гибриды крыс и мышей. Успехи в развитии генетики человека сделали возможными предупреждение и лечение наследственных заболеваний. Практические мероприятия, направленные на поддержание наследственного здоровья человека, на охрану генофонда человечества, осуществляются через систему медико-генетических консультаций. Их основная цель — информировать заинтересованных лиц о вероятности риска появления в потомстве больных.Современная генетика предлагает возможность создавать новые организмы для тех или иных целей, поставленных человеком: растения, синтезирующие удобрения прямо из воздуха; бактерии, производящие человеческие белки; бактерии, которые питаются загрязняющими веществами или производящие белки из нефти; вирусы, переносящие человеческие гены Перспективы развития генетики С каждым годом мы становимся чуть ближе к тому, чтобы предотвращать развитие наследственных болезней на этапе развития плода. Основной инструмент, способный на такое волшебство, — это генное редактирование при помощи технологии CRISPR. Именно она позволяет буквально влезать в ДНК, удалять или преобразовывать нужные гены. По прогнозам генетиков, уже к концу второго десятилетия XXI века на смену привычным прививкам придут генетические вакцины, и медики получат возможность навсегда покончить с такими неизлечимыми болезнями, как рак, болезнь Альцгеймера, диабет, астма. В этом направлении ведутся научные исследования, которое имеет свое название - генотерапия. По некоторым прогнозам примерно в 2020 году на свет будут появляться исключительно здоровые дети: уже на эмбриональной стадии развития плода генетики смогут исправлять наследственные неполадки. Ученые прогнозируют, что в 2050 году будут попытки по усовершенствованию человеческого вида. Мы прочтем всё, что записано в наших хромосомах, и научимся это понимать, воспользуемся этим, чтобы исправить все найденные ошибки. К этому времени научимся проектировать людей определенной специализации: математиков, физиков, художников, поэтов, а может быть, и гениев. Исполнится мечта человека: процессом старения, несомненно, можно будет управлять, а там недалеко и до бессмертия План: 1 Введение 2 Успехи генетики 3 Перспектива современной генетики Список литературы / References 1. Курчанов Н.А. Генетика человека с основами общей генетики. «Учебное пособие»: СпецЛит; Санкт-Петербург, 2009. 122 с. 2. ГриневВ.В. Генетика человека: курс лекций / В.В. Гринев. Минск: БГУ, 2006. 131 с. 3. ДуменкоВ.Б. О здоровье тела, разума, души. Справочник. К. Гиперион, 2014. 464 с. 4. БабаевМ.Ш., Мамедова Р.Ф. История развития биологии. С. 381. 5. Багрикова С.В. Основные положения современной генетики. Генная инженерия, 2017. [Электронный ресурс]. Режим доступа: http://stomatologo.ru/s-v-bagrikova.html? page=28. С. 38/. 6. КушнировВ.В. Генетика человека: состояние и перспективы. [Электронный ресурс]. Режим доступа::http://mygenome.su/articles/100/ Мой геном, научно-популярный портал по генетике/ 7. Мир знаний, перспективы развития генетики. С. 3. [Электронный ресурс]. Режим доступа: http:// mirznanii.eom/a/151509-3/perspektivy-razvitiya-genetiki-3. Медицина и здоровье 8. Генетика и человек - Достижения и проблемы современной генетики. [Электронный ресурс]. Режим доступа: http://1doklad.ru/index.php?catid=9%3A2010-06-30-10-37-48&id= 106%3A2010-07-28-11 -25-20&Itemid =11&option= com_content& view =article&limitstart=6- с. 9. Возможности современной генетики. [Электронный ресурс]. Режим доступа: http:// biofile.ru/bio/ 5240. html/. Мейоз и его биологическое значениеОпределение Мейоз (редукционное деление клетки) — деление, в процессе которого из одной диплоидной (2n) клетки получаются 4 гаплоидные (n) клетки. Мейоз и его биологическое значениеОпределение Мейоз (редукционное деление клетки) — деление, в процессе которого из одной диплоидной (2n) клетки получаются 4 гаплоидные (n) клетки. Так как у дочерних клеток происходит уменьшение (редукция) числа хромосом с 2n до n, такое деление названо редукционным. Схема мейоза  Мейоз у животных наблюдается при формировании гамет (гаметогенезе). Мейоз у растений и грибов, как правило, происходит при образовании гаплоидных спор. У различных одноклеточных эукариот мейоз может наблюдаться на разных стадиях жизненного цикла. Для восстановления диплоидности в цикле всегда необходимо слияние гаплоидных клеток (оплодотворение). Мейоз состоит из двух делений. Первое из них является собственно редукционным, то есть именно в ходе первого деления уменьшается плоидность клетки. Причиной этого служит расхождение гомологичных хромосом («материнской» и «отцовской») по двум разным дочерним клеткам. Второе деление аналогично митозу и называется эквационным (то есть «равным»). Плоидность в результате второго деления не меняется. В ходе этого деления, как и при митозе, расходятся сестринские хроматиды (копии ДНК). Между двумя делениями мейоза отсутствует репликация ДНК (так как «цель» мейоза — уменьшить плоидность клетки, увеличивать количество ДНК здесь незачем). В профазе I деления мейоза происходит важнейший процесс, относящийся к генетической рекомбинации — кроссинговер, то есть обмен участками гомологичных хромосом. В результате этого процесса создаются новые комбинации генов в потомстве. Хромосомы как целое не передаются напрямую от бабушек и дедушек внукам, а «реконструируются» в каждом поколении в процессе кроссинговера. В нижеследующей таблице приведено описание фаз мейоза в клетке, для которой n=2, 2n=4. В каждом наборе есть три хромосомы, различающиеся по размеру. Материнский и отцовский хромосомные наборы выделены синим и красным. Протекание мейоза, как правило, нарушается в клетках гибридных организмов, т. к. в профазе I должно происходить попарное слияние (конъюгация) гомологичных хромосом, а у гибридов набор материнских генов не гомологичен отцовскому. Данный механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и в конечном счете к нежизнеспособности гамет, а следовательно, к стерильности (бесплодию) гибридов. В селекции для преодоления стерильности гибридов искусственно вызывают полиплоидность (кратное увеличение) хромосомных наборов. В этом случае каждая хромосома конъюгирует с соответствующей хромосомой своего набора. Значение мейозаПоловые клетки родителей, образовавшиеся путем мейоза, обладают гаплоидным набором (n) хромосом. В зиготе при объединении двух таких наборов число хромосом становится диплоидным (2n). Формирование нового организма происходит путем митотических делений зиготы, и каждая его клетка содержит диплоидный (2n) набор хромосом. Каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому. Исходя из этого: Мейоз является основой комбинативной изменчивости благодаря кроссинговеру (профаза I) и независимому расхождению гомологичных хромосом (анафаза I и II). Благодаря уменьшению количества хромосом в гаметах в новых организмах поддерживается постоянный диплоидный (2n) набор хромосом. План 1 Определение 2 Процесс мейоза 3 Значение мейоза МЕХАНИЗМЫ РЕКОМБИНАЦИИРекомбинация генов осуществляется различными способами. Этот процесс может быть связан с перераспределением целых хромосом. Такой механизм в соответствии с третьим законом Г. Менделя обеспечивает независимое наследование несцепленных генов и признаков. Чаще всего рекомбинацию в узком смысле слова связывают с кроссинговером, т. е. с пе- рекомбинацией генов, локализованных в гомологичных хромосомах. Цитологическая демонстрация кроссинговераВ профазе I мейоза на стадии диплотены у многих организмов хорошо различимы характерные фигуры, образуемые гомологичными хромосомами, - хиазмы. В 1909 г. Ф. Янссене предположил, чгго образование хиазм связано с обменами гомологичными участками гомологичных хромосом. Позднее Т.Х. Морган связывай хиазмы с кроссинговером, происходящим согласно гипотезе К. Бриджеса (1915) по механизму разрыв-воссоединение. Дарлингтон, исходя из числа хиазм в пахитене, определил общую длину каждой из хромосом кукурузы. При этом он принял, что одна хиазма образуется на участке в 50 сМ. Полученный на основании цитологических данных результат хорошо согласовался с длинами групп сцепления, вычисленными обычным способом исходя из частот кроссинговера при гибридологическом анализе. Параллелизм событий, регистрируемых генетически и цитологически при рекомбинации сцепленных генов, был продемонстрирован в 1931 г. для двух объектов: кукурузы и дрозофилы. В обоих случаях был использован принцип: сопоставление результатов кроссинговера с физическими обменами гомологичных участков гомологичных хромосом. Очевидно, для такой работы скрещиваемые формы должны быть дигетеро- зиготны как по генетическим, так и по цитологическим маркерам одной пары хромосом. Харриет Крейтон и Барбара МакКлинток решили эту проблему относительно кукурузы. Была исследована форма, дигетерозиготная по генам: с (colorless) - бесцветный алероновый слой эндосперма, рецессив; с+ - окрашенный, доминант; wx (yvaxy) - эндосперм, а также пыльцевые зерна, которые содержат амилопектин, вследствие чего окрашиваются йодом в красный цвет, рецессив; wx - эндосперм и пыльцевые зерна, содержащие амилозу, окрашиваемую йодом в синий цвет, доминант. Дарлингтон, исходя из числа хиазм в пахитене, определил общую длину каждой из хромосом кукурузы. При этом он принял, что одна хиазма образуется на участке в 50 сМ. Полученный на основании цитологических данных результат хорошо согласовался с длинами групп сцепления, вычисленными обычным способом исходя из частот кроссинговера при гибридологическом анализе. Параллелизм событий, регистрируемых генетически и цитологически при рекомбинации сцепленных генов, был продемонстрирован в 1931 г. для двух объектов: кукурузы и дрозофилы. В обоих случаях был использован принцип: сопоставление результатов кроссинговера с физическими обменами гомологичных участков гомологичных хромосом. Очевидно, для такой работы скрещиваемые формы должны быть дигетеро- зиготны как по генетическим, так и по цитологическим маркерам одной пары хромосом. Харриет Крейтон и Барбара МакКлинток решили эту проблему относительно кукурузы. Была исследована форма, дигетерозиготная по генам: с (colorless) - бесцветный алероновый слой эндосперма, рецессив; с+ - окрашенный, доминант; wx (yvaxy) - эндосперм, а также пыльцевые зерна, которые содержат амилопектин, вследствие чего окрашиваются йодом в красный цвет, рецессив; wx - эндосперм и пыльцевые зерна, содержащие амилозу, окрашиваемую йодом в синий цвет, доминант. Эти же самки несли гетероморфные Х-хромосомы: одну сильно укороченную вследствие транслокации ее бесцентромерного участка на четвертую микрохромосому, другую - Г-образную вследствие транслокации на нее фрагмента У-хромосомы. Таких самок скрещивали с самцами, несущими рецессивные аллели исследуемых генов (сг, В+) в Х-хромосоме обычной формы и имевшими обычную У-хромосому. Результат кроссинговера у самок в таком скрещивании - рекомбинантные классы - изучали цитологически. Для этой цели брали только самок, поскольку самцы имеют субметацентрическую, т. е. разноплечую У-хромосому, которую можно спутать с Г-образной ^-хромосомой. Все четыре класса самок-потомков отличались друг от друга по фенотипу: 1) нерекомбинантные: 2) рекомбинантные: 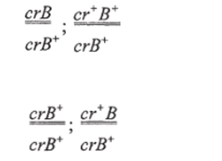 а также по структуре своих X-хромосом. Исследовав 374 самки, К. Штерн показал, что кроссинговер между генами сг и В сопровождался физическим обменом между Л'-хромосомами. Эксперименты X. Крейтон и Б. МакКлинток с кукурузой, К. Штерна с дрозофилой доказали, что в основе кроссинговера лежит реальный обмен участками гомологичных хромосом, однако механизм этого обмена остался неясным. Наряду с гипотезой К. Бриджеса (механизм разрыв-слияние) рассматривался и другой механизм, предложенный Дж. Беллингом (1933), изучавшим мейоз у растений. Согласно этой гипотезе, разрывов и перевоссоединения хромосом не происходит, а кроссинговер приурочен к стадии воспроизведения хромосом. При этом, как считал Дж. Беллинг, сначала воспроизводятся хромомеры, затем их соединяют хромонемы. Такое соединение может привести к рекомбинантным сочетаниям хромомеров. В 1930 г. X. Винклером была предложена еще одна гипотеза, согласно которой в потомстве дигетерозиготы рекомбинантные классы могут появляться вследствие направленных изменений аллелей под влиянием друг друга - гипотеза конверсии: М 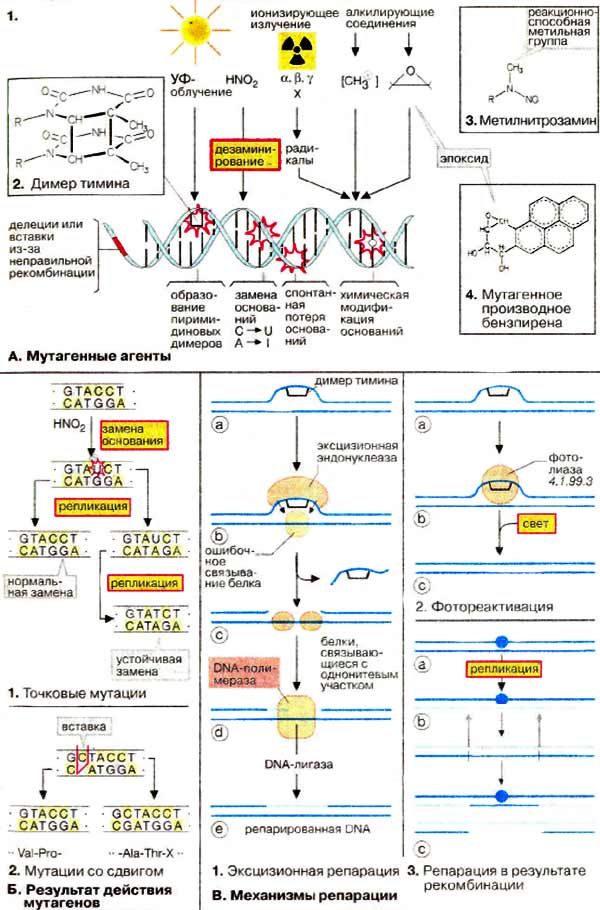 утация и репарация утация и репарацияГенетическая информация кодируется последовательностью оснований ДНК и поэтому изменения в структуре или последовательности азотистых оснований приводят к мутациям. Многие мутагены вызывают нарушения регуляции роста и деления клеток и поэтому являются канцерогенными. Изменение в структуре генов (мутация) — важный фактор биологической эволюции. В то же время слишком высокая скорость мутаций ставит под вопрос существование индивидуальных организмов или целых видов. Поэтому клетки обладают механизмами восстановления (репарации), которые корректируют большинство изменений ДНК, вызываемых мутациями. А. Мутагенные агенты Мутации могут возникать в результате либо воздействия физических или химических факторов, либо ошибок в процессе репликации и рекомбинации ДНК. Одним из наиболее важных физических мутагенов является ионизирующая радиация. Она приводит к образованию в клетке свободных радикалов (молекул с неспаренными электронами, см. с. 20), которые исключительно реакционноспособны и могут повредить ДНК. Коротковолновый ультрафиолетовый свет (УФ) также оказывает мутагенное действие, особенно на клетки кожи. Наиболее распространенным химическим изменением, вызванным ультрафиолетовым облучением, является образование тиминовых димеров, когда два соседних тиминовых оснований ковалентно связываются друг с другом. Это приводит к ошибкам при считывании ДНК во время репликации и транскрипции. Из множества химических мутагенов здесь приведен только несколько примеров. Азотистая кислота (HNO2) и гидроксиламин (NH2OH) дезаминируют азотистые основания, т. е. превращают цитозин в урацил, а аденин в инозин. Алкилирующие соединения имеют реакционные группы, которые могут образовывать ковалентные связи с азотистыми основаниями, входящими в ДНК. Метилнитрозамины распадаются с образованием реакционноспособного метилкатиона (СН3-), который метилирует группы ОН и NH2 в ДНК. Ароматический углеводород бензпирен сам по себе безвреден, но в результате метаболической трансформации образует производные, обладающие канцерогенным действием. За счет гидроксилирования одного из колец он превращается в реакционноспособный эпоксид, который алкилирует аминогруппу гуанина и других азотистых оснований. Токсичен и свободный радикал бензпирена. Б. Результат действия мутагенов Азотистая кислота вызывает точковые мутации. Например, С превращается в U, который в следующем цикле репликации образует пару с А (вместо G). после чего изменение принимает необратимый характер. Мутации типа вставки или выпадения некоторого числа нуклеотидов, не кратного трем, ведут к ошибочной трансляции всей ДНК, поскольку они сдвигают рамку считывания (2). При трансляции измененная мРНК будет интерпретироваться рибосомами по-другому, приводя к совершенно иной аминокислотной последовательности, что показано на простом примере (2). В. Механизмы репарации Важным механизмом удаления повреждений в ДНК является эксцизионная репарация. Специфическая нуклеаза удаляет небольшой сегмент ДНК, включающий поврежденный участок. Удаленный участок восстанавливается ДНК-полимеразой, использующей в качестве матрицы комплементарную цепь. Наконец, оставшийся одноцепочечный разрыв закрывается ДНК-лигазой. Тиминовые димеры могут быть удалены фотореактивацией. Специфическая фотолиаза связывается с дефектным участком ДНК и после облучения расщепляет димер с образованием отдельных нуклеиновых оснований. Третий механизм — это репарация в результате рекомбинации (3, показано в упрощенном виде). В этом процессе участок, содержащий повреждение, пропускается во время репликации. Образующаяся брешь закрывается путем сдвига соответствующего сегмента из правильно реплицированной второй цепи. Новая брешь ликвидируется с участием полимераз и ДНК-лигаз. В завершение первоначальный дефект устраняется путем вырезания. План 1 Мутагенные агенты 2Результат действия мутагенов 3 Механизмы репараций План Введение Понятие «ген». Развитие представлений о нем Структура генов. Классификация генов Функции генов Список использованной литературы. Введение Цель моего реферата – попытка раскрыть фундаментального понятия современной генетики – понятия «ген». Генетика – молодая наука, она начала развиваться только в XX веке. Генетика изучает законы двух фундаментальных свойств живых организмов - наследственности и изменчивости, лежащие в основе эволюции органического мира и деятельности человека по созданию новых сортов культурных растений, пород животных и штаммов микроорганизмов.Наследственность – это свойство организма передавать свои признаки и особенности развития следующим поколениям.Изменчивость – это свойство организмов приобретать новые признаки в процессе индивидуального развития.Оба этих фундаментальных свойств организмов (и наследственность, и изменчивость) осуществляются генами. Гены хранят и передают информацию об организме последующим поколениям. 1. Понятие «ген». Развитие представлений о нем Ген – функциональная единица наследственного материала. Ген (от греч. genos — род, происхождение) – участок молекулы геномной нуклеиновой кислоты, характеризуемый специфической для него последовательностью нуклеотидов, представляющий единицу функции, отличной от функций других генов, и способный изменяться путем мутирования. 2. Структура генов Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар нуклеотидов, в которых закодирована генетическая информация о первичной структуре белка (число и последовательность аминокислот). Для регулярного правильного считывания информации в гене должны присутствовать: кодон инициации, множество смысловых кодонов и кодон терминации. Три подряд расположенных нуклеотида представляют собой кодон, который и определяет, какая аминокислота будет располагаться в данной позиции в белке. Например, в молекуле ДНК последовательность оснований ТАС является кодоном для аминокислоты метионина, а последовательность ТТТ кодирует фенилаланин. В молекуле иРНК вместо тимина (Т) присутствует основание урацил (У). Таблица генетического кода во всех руководствах представлена именно символами иРНК. Из 64 возможных кодонов смысловыми являются 61, а три триплета — УАА, УАГ, УГА — не кодируют аминокислоты и поэтому были названы бессмысленными, однако на самом деле они представляют собой знаки терминации трансляции.Для прокариот характерна относительно простая структура генов. Так, структурный ген бактерии, фага или вируса, как правило, контролирует одну ферментативную реакцию. Специфичным для прокариот является оперонная система организации нескольких генов. Гены одного оперона (участка генетического материала, состоящего из 1, 2 и более сцепленных структурных генов, которые кодируют белки (ферменты), осуществляющие последовательные этапы биосинтеза какого-либо метаболита; в оперон эукариот входит, как правило, 1 структурный ген; оперон содержит регуляторные элементы) расположены в кольцевой хромосоме бактерии рядом и контролируют ферменты, осуществляющие последовательные или близкие реакции синтеза (лактозный, гистидиновый и др. опероны). 3. Классификация генов Накопленные знания о структуре, функциях, характере взаимодействия, экспрессии, мутабильности и других свойствах генов породили несколько вариантов классификации генов.По месту локализации генов в структурах клетки различают расположенные в хромосомах ядра ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями.По функциональному значению различают структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены — последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.).По влиянию на физиологические процессы в клетке различают летальные, условно летальные, супервитальные гены, гены-мутаторы, гены-антимутаторы и др.Следует отметить, что любые биохимические и биологические процессы в организме находятся под генным контролем. Так, деление клеток (митоз, мейоз) контролируется несколькими десятками генов; группы генов осуществляют контроль восстановления генетических повреждений ДНК (репарация). Онкогены и гены — супрессоры опухолей участвуют в процессах нормального деления клеток. Индивидуальное развитие организма (онтогенез) контролируется многими сотнями генов. Мутации в генах приводят к измененному синтезу белковых продуктов и нарушению биохимических или физиологических процессов. 4. Функции генов В процессе реализации наследственной информации, заключенной в гене, проявляется целый ряд его свойств. Определяя возможность развития отдельного качества, присущего данной клетке или организму, ген характеризуется дискретностью действия (от лат. discretus — разделенный, прерывистый), прерывностью (интроны и экзоны). Дискретность наследственного материала, предположение о которой высказал еще Г. Мендель, подразумевает делимость его на части, являющиеся элементарными единицами, - гены. В настоящее время ген рассматривают как единицу генетической функции. Он представляет собой минимальное количество наследственного материала, которое необходимо для синтеза тРНК, рРНК или полипептида с определенными свойствами. Ген несет ответственность за формирование и передачу по наследству отдельного признака или свойства клеток, организмов данного вида. Кроме того, изменение структуры гена, возникающее в разных его участках, в конечном итоге приводит к изменению соответствующего элементарного признака.Ввиду того что в гене заключается информация об аминокислотной последовательности определенного полипептида, его действие является специфичным. Однако в некоторых случаях одна и та же нуклеотидная последовательность может детерминировать синтез не одного, а нескольких полипептидов. Это наблюдается в случае альтернативного сплайсинга у эукариот и при перекрывании генов у фагов и прокариот. Очевидно, такую способность следует оценить как множественное, или плейотропное, действие гена (хотя традиционно под плейотропным действием гена принято понимать участие его продукта – полипептида – в разных биохимических процессах, имеющих отношение к формированию различных сложных признаковП 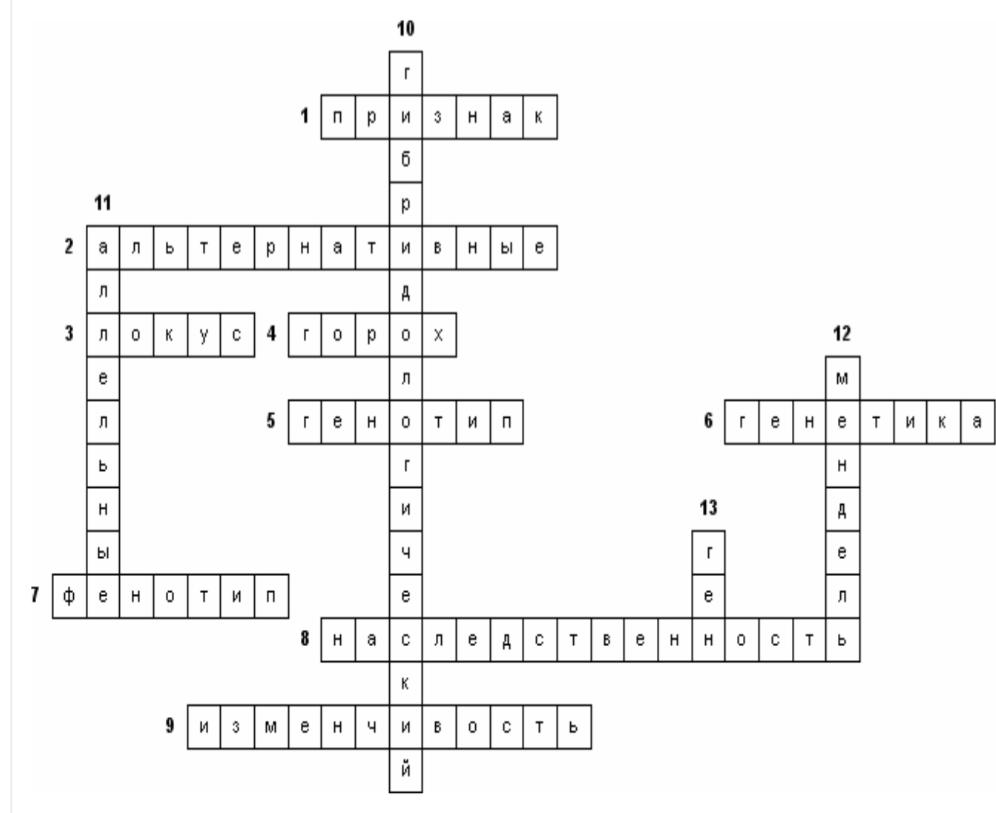 о горизонтали 1.Свойство, определяемое геном. 2.Контрастные (взаимоисключающие) признаки. 3.Место в хромосоме, где располагается ген ответственный за развитие данного признака. 4.Растение, на котором Г. Мендель сделал основные свои открытия. 5.Совокупность всех генов одного организма. 6.Наука о закономерностях наследования признаков у организмов. 7.Совокупность всех признаков организма. 8.Способность родителей передавать свои признаки, свойства и особенности развития следующему поколению. 9.Способность организмов, приобретать новые признаки и свойства. По вертикали 10.Метод, предложенный Г.Менделем, при котором скрещивающиеся организмы различаются по одной паре взаимоисключающих признаков. 11.Гены, расположенные в одних и тех же локусах гомологичных хромосом и ответственных за развитие одного признака. 12.Чешский ученый, основоположник науки генетики. 13. Участок молекулы ДНК, определяющий возможность развития отдельного элементарного признака, или синтез одной белковой молекулы. |