Вопрос 1 Предмет и задачи генетики. Уровни организации наследственного материала, их характеристика
 Скачать 149.67 Kb. Скачать 149.67 Kb.
|

|
Вопрос 14 и 15 Эволюция понятия гена. Взгляды Н.Кольцова на биохимическую структуру гена. Экспериментальные доказательства роли ДНК в передаче наследственной информации ( явление трансформации в опытах Гриффитса, трансдукции, эксперименты Фринкель-Конрата с вирусом табачной мозайки, опыт Херши и Чейз с бактериофагом 80-е гг. XIX в. ознаменовались важными достижениями в области цитологии: были описаны митоз и мейоз — деление соответственно соматических и половых клеток, в ходе которых закономерно между дочерними клетками распределяются ядерные структуры —хромосомы (В. Вольдейер, 1888). Данные о характере распределения хромосом в процессе клеточного деления позволили в начале XX в. Т. Бовери (1902—1907) и У. Сетгону (1902—1903) сделать вывод о том, что преемственность свойств в ряду поколений клеток и организмов определяется преемственностью их хромосом. Хромосомы стали рассматривать как материальные носители наследственной программы. Дальнейшая разработка хромосомной теории наследственности, объединяющей представления о наследственных задатках и хромосомах, была осуществлена в начале XX в. Т. Морганом и его сотрудниками. В опытах, выполненных на дрозофиле, было подтверждено ранее высказанное предположение о роли хромосом в обеспечении наследственности. Установлено, что гены размещаются в хромосомах, располагаясь в них в линейном порядке. Гены каждой хромосомы образуют группу сцепления, число которых определяется количеством хромосом в половых клетках. Гены одной группы сцепления наследуются, как правило, совместно. Однако в ряде случаев происходит их перекомбинация в связи с кроссинговером (см. разд. 5.3.2), частота которого зависит от расстояния между генами. Таким образом, в хромосомной теории нашел отражение один из важнейших принципов генетики — единство дискретности и непрерывности наследственного материала. Необходимо отметить, что также в начале XX в. были обнаружены факты, которые доказывали наличие в клетках внехромосомного наследственного материала, располагающегося в различных цитоплазматических структурах и определяющего особую цитоплазматическую наследственность (К. Корренс, 1908). Примерно в это же время X. де Фризом (1901) были заложены основы учения о мутационной изменчивости, связанной с внезапно возникающими изменениями в наследственных задатках или хромосомах, что приводит к изменениям тех или иных признаков организма. В последующие годы было обнаружено мутагенное действие на хромосомы и гены рентгеновских лучей, радиационного излучения, определенных химических веществ и биологических агентов. В результате этих исследований стало очевидным, что наследственность и изменчивость обусловлены функционированием одного и того же материального субстрата. В первые десятилетия XX в. были получены данные, свидетельствующие в пользу зависимости состояния признаков от характера взаимодействия генов, что выходило за рамки отношений доминантности и рецессивности, описанных еще Менделем. Отсюда появилось представление о генетическом аппарате как о системе взаимодействующих генов — генотипе, который сосредоточен в хромосомном наборе — кариотипе. Изучение химического состава хромосом выявило два основных вида соединений, образующих эти структуры,— белки и нуклеиновые кислоты. В первой половине XX в. исследователями решался вопрос о химической природе субстрата наследственности и изменчивости. Первоначально высказывались предположения в пользу белков. В 1928 г. Ф. Гриффитом был поставлен опыт на пневмококках, в котором наблюдалось изменение (трансформация) некоторых наследственных свойств одного бактериального штамма под влиянием материала, полученного из убитых клеток другого штамма. Химическая природа вещества, трансформирующего наследственные свойства бактерий, была установлена лишь в 1944 г. О. Эйвери, доказавшим его принадлежность к нуклеиновым кислотам (ДНК). Другими доказательствами участия ДНК в обеспечении наследственности и изменчивости являются: 1) постоянство содержания ДНК во всех типах соматических клеток организма; 2) соответствие содержания ДНК плоидности клеток (в соматических клетках ее вдвое больше, чем в половых, в полиплоидных клетках оно соответствует количеству наборов хромосом); 3) явление генетической рекомбинации у бактерий при их конъюгации, в ходе которой осуществляется проникновение части ДНК из одной клетки в другую и изменение свойств последней; 4) изменение наследственных свойств бактериальных клеток путем переноса ДНК от одного штамма к другому с помощью ДНК-фага — явление трансдукции; 5) инфицирующая активность изолированной нуклеиновой кислоты вирусов. Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). В этом вирусе содержится РНК, а не ДНК. Было известно, что разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком. Следовательно, не только ДНК, но и РНК может служить носителем генетической информации. Фаги (бактериофаги) - это вирусы, размножающиеся в бактериях. Суть опыта: фаги, у которых белковая оболочка была мечена радиоактивной серой (S35), а ДНК - радиоактивным фосфором (Р32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р32, а в бактериях - S35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК. Отсюда следовал однозначный вывод о том, что именно ДНК выполняет генетическую функцию - несет информацию как о создании новых копий ДНК, так и о синтезе фаговых белков 1957г. Вопрос 16 Молекулярное строение генов прокариот и эукариот. Концепция гена «цистрона» для прокариот и «мозаичного» для эукариот. Клеткам прокариотического типа свойственны малые размеры (не более 0,5—3,0 мкм в диаметре или по длине), отсутствие обособленного ядра, так что генетический материал в виде ДНК не отграничен от цитоплазмы оболочкой. В клетке отсутствует развитая система мембран. Генетический аппарат представлен ДНК единственной кольцевой хромосомы, которая лишена основных белков — гистонов (гистоны являются белками клеточных ядер). Благодаря значительному количеству диаминокислот аргинина и лизина гистоны имеют щелочной характер. Цистрон — наименьшая единица генетической экспрессии, кодирующая одну субъединицу белковой молекулы. Поэтому вышеупомянутую формулу можно перефразировать как «один цистрон — одна субъединица". Строение мозаичного гена В конце 70-х годов было выяснено, что у эукариот имеются гены, которые содержат «лишнюю» ДНК, не представленную в молекуле мРНК. Они получили название мозаичных, прерывистых генов; генов, имеющих экзон-интронное строение. 1.Мозаичные гены эукариот имеют больший размер, чем последовательность нуклеотидов, представленная в мРНК (3-5%). 2.Мозаичные гены состоят из экзонов и интронов. Интроны удаляются из первичного транскрипта и отсутствуют в зрелой мРНК, которая состоит только из экзонов. Число и размеры интронов и экзонов индивидуальны для каждого гена, но интроны по размерам значительно больше экзонов. 4. Порядок расположения экзонов в гене совпадает с их расположением в мРНК. 5.На границе экзон-интрон имеется определённая постоянная последовательность нуклеотидов (ГТ - АГ), присутствующая во всех мозаичных генах. 6. Экзон одного гена может быть интроном другого. 7. В мозаичном гене иногда нет однозначного соответствия между геном и кодируемым им белком, то есть одна и та же последовательность ДНК может принимать участие в синтезе различных вариантов белка. 9. Особенности строения мозаичного гена позволяют осуществлять альтернативный сплайсинг (экзон L - экзон 2,3 или экзон S - экзон 2,3): синтезировать несколько вариантов белка на основе информации одного гена; создавать удачные комбинации белков, а если таковые неудачны, то производить отбор на уровне мРНК при сохранении неизменной ДНК. Вопрос 17 Ген - функциональная единица наследственности. Гипотеза «один ген- один фрагмент» и ее развитие в концепцию «один цистрон – один полипептид». Центральная догма биологии. Ген/цистрон– участок ДНК, имеющий определенную последовательность нуклеотидов, содержащий информацию о структуре какого-либо одного белка. Ген - функциональная единица наследственной информации. Концепция, согласно которой одним геном может кодироваться только один фермент; более строго это соотношение отражено в теории “один ген - один полипептид”, т.к. один фермент может быть гетерополимером и включать полипептидные цепи, кодируемые разными генами. Один ген молекулы ДНК кодирует один белок, отвечающий за одну химическую реакцию в клетке. Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении.. Вопрос 18 Требования, предъявляемые к материальному субстрату, ответственному за несение генетической информации. Цистрон-регулятор, цистрон-оператор, структурные цистроны. Этапы транскрипции и трансляции Требования, предъявляемые к субстрату наследственности: Способность к самовоспроизведению – вещество должно обеспечить преемственность свойств в поколениях. Уникальность – вещество должно иметь структуру, объясняющую существование миллионов видов и неповторимость. Специфичность – структура вещества должна предполагать синтез специфических белков. Цистрон-регулятор – ген, контролирующий синтез белка-репрессора, родственного к структуре оператора. Цистрон- оператор – функциональный ген, расположенный в начале оперона, включающий и выключающий структуры генов. Структурные цистроны – расположены рядом и образуют оперон. Они программируют синтез ферментов, участвующих в последовательно идущих ферментативных реакциях одного метаболического цикла. Транскрип́ ция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы. 1)Инициация – первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором и образование первой межнуклеотидной связи. 2)Элонгация – последовательное удлинение растущей цепи РНК. 3)Терминация (окончание транскрипции). Трансляция - перевод генетической информации с нуклеотидного кода, записанного в молекулах мРНК, в определенную последовательность аминокислот в полипептидной цепи синтезируемого белка. Этапы трансляции - инициация (начало синтеза), элонгация (удлинение, наращивание полипептидной цепи), терминация (окончание синтеза). Вопрос 19 Общие принципы генетического контроля экспрессии генов. Роль генетических факторов в регуляции генной активности. Общие принципы генетического контроля экспрессии генов. Важнейшим фактором регуляции генной активности являются элементы генома, отвечающие за синтез регуляторных белков,— гены-регуляторы. Соединяясь с определенными нуклеотидными последовательностями ДНК, предшествующими структурной части регулируемого гена,—операторами, белки-регуляторы способствуют или препятствуют соединению РНК-полимеразы с промотором. Если белок-регулятор взаимодействует с оператором, занимающим часть промотора или расположенным между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соединиться с промоторной последовательностью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществляется негативный контроль экспрессии гена со стороны гена-регулятора. Если промотор обладает слабой способностью соединяться с РНК-полимеразой, а ему предшествует область, узнаваемая белком-регулятором, присоединение последнего непосредственно перед промотором к молекуле ДНК облегчает связывание РНК-полимеразы с промотором, вслед за чем следует транскрипция. Такие белки называют активаторами (или апоиндукторами), а контроль экспрессии гена со стороны гена-регулятора — позитивным. Роль негенетических факторов в регуляции генной активности. Наряду с генетическими факторами в регуляции экспрессии генов на стадии транскрипции принимают участие негенетические факторы — эффекторы. К ним относят вещества небелковой природы, взаимодействующие с белками-регуляторами и изменяющие их способность соединяться с нуклеотидными последовательностями операторов. В зависимости от результатов такого взаимодействия среди эффекторов различают индукторы, запускающие транскрипцию, икорепрессоры, препятствующие ей.Индукторы могут инактивировать белки-репрессоры, которые перестают соединяться с операторами, или повышать способность белков-активаторов (апоиндукторов) к связыванию с ними, что облегчает соединение РНК-полимеразы с промотором. В результате такого воздействия на регуляторные белки регулируемые гены активно транскрибируются.Корепрессоры могут модифицировать апоиндукторы, теряющие при этом способность соединяться с операторами, или активировать репрессоры, находящиеся в неактивном состоянии. Следствием такого взаимодействия эффектора с белками-регуляторами является невозможность соединения РНК-полимеразы с промотором и отсутствие транскрипции. Вопрос 20 и 21 Изменчивость и ее формы. Понятие о фенотипической изменчивости. Фенокопия. Комбинативная изменчивость, ее формы и биологическая роль Изменчивость – это универсальное свойство живых организмов приобретать новые признаки под действием среды (как внешней, так и внутренней). Выделяют два типа изменчивости: ненаследственную(модификационную, фенотипическую) и наследственную(генотипическую).Модификационная (фенотипическая) изменчивость заключается в том, что под действием разных условий внешней среды у организмов одного вида, генотипически одинаковых, наблюдается изменение признаков (фенотипа). Фенокопии — изменения фенотипа под влиянием неблагоприятных факторов среды, по проявлению похожие на мутации. 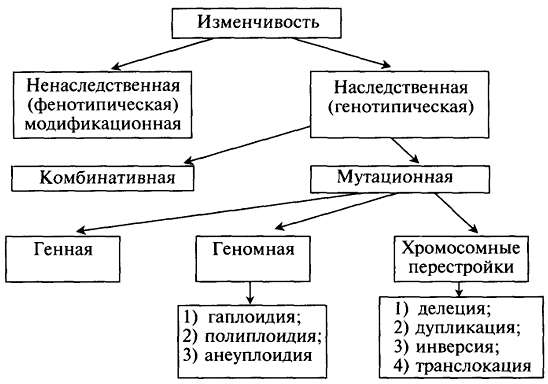 Комбинативная изменчивость, являясь результатом перекомбинирования генов родительских особей в генотипах потомков, основывается на трёх основных механизмах. +1. Независимое расхождение в дочерние клетки (сперматоциты II, ооцит II и первое редукционное тельце) гомологичных хромосом из каждой пары (имеет место при I делении мейоза в ходе гаметогенеза). +2. Случайное сочетание гамет, а следовательно, гомологичных (отцовской и материнской) хромосом при оплодотворении. +3. Обмен отдельными аллелями между гомологичными хромосомами в процессе кроссинговера мейоза. Вопрос 22 Понятие о мутационной изменчивости. Мутации в зависимости от места возникновения (соматические, генеративные), значение. Примеры, основные положения мутационной теории. Мутационная изменчивость связана с изменениями последовательности нуклеотидов в молекулах ДНК, выпадения и вставок крупных участков в молекулах ДНК, изменений числа молекул ДНК (хромосом). Сами подобные изменения называются мутациями .Мутации наследуются. СОМАТИЧЕСКИЕ МУТАЦИИ Мутации различного ранга возникающие в соматических клетках организма, наследуются потомками этих клеток и делают организм мозаиком, т.е. особью со смешанными популяциями клеток. ГЕНЕРАТИВНЫЕ МУТАЦИИ Изменения наследственной программы половых клеток человека приводят к рождению потомства с различными наследственно обусловленными болезнями, в зависимости от ранга мутаций — генными или хромосомными. Различные генные мутации по-разному сказываются на жизнеспособности организма, причем в случае их рецессивности они могут долго не проявляться фенотипически у потомков. Хромосомные перестройки и геномные мутации приводят к выраженным отклонениям в развитии и часто являются причиной гибели организма на разных стадиях его онтогенеза, обычно в раннем эмбриогенезе. Основные положения мутационной теории. 1.Мутации возникают внезапно, без переходов, скачкообразно 2.Новые изменения константны, устойчивы, наследуются. 3.Мутации не образуют вариационных рядов, т.е. они являются качественными, а не количественными. 4.Мутации возникают в разных направлениях, могут быть как полезными, так и вредными, т.е. они не носят адаптивного характера. 5.Выявление мутаций зависит от количества проанализированных особей. 6.Одни и те же мутации могут возникать повторно. 7.Мутации дают начало новым видам. 8.В эволюции большее значение имеют множественные мелкие мутации, а не резкие крупные изменения. 9.Мутации автогенны, т. е. самопроизвольны. Вопрос 23 Мутации в зависимости от причины возникновения (спонтанные,индуцированные), значение, примеры. Мутагенные факторы. Канцерогенез. Индуцированные мутации – вызваны специально направленными воздействиями, повышающими мутационный процесс. Спонтанные мутации – возникают под влияние неизвестных природных факторов, повышающих мутационный процесс, чаще всего как результат ошибок при репликации ДНК. Мутагенным действием обладают факторы физической, химической и биологической природы. Мутагенные факторы среды – факторы, вызывающие появление мутаций. Физические: Ионизирующая радиация – рентгеновские лучи, α-, β-, γ-лучи Химические: Неорганические и органические соединения, например кислоты, щелочи, перекиси, соли металлов, формальдегид, пестициды, дефолианты, гербициды, колхицин и др. Некоторые вещества способны усиливать мутационный эффект в сотни раз по сравнению со спонтанным. Их называют супермутагенами. К ним относят нитрозосоединения – иприт, диэтилнитрозамин, уретан и др. Некоторые лекарственные препараты также обладают мутагенным эффектом, например, цитостатики, производные этиленимина, нитрозомочевина. Они повреждают ДНК в процессе репликации. Биологической природы: Вирусы оспы, кори, ветряной оспы, эпидемического паротита, гепатита, краснухи и др. способны вызывать разрывы хромосом. Вирусы могут усиливать темпы мутации клеток хозяина за счет подавления активности репарационных систем. Канцерогенез — сложный патофизиологический процесс зарождения и развития опухоли. |