Экзамен по биохимии 2 курс. Вопросы итоговой аттестации по биохимии
 Скачать 1.2 Mb. Скачать 1.2 Mb.
|

|
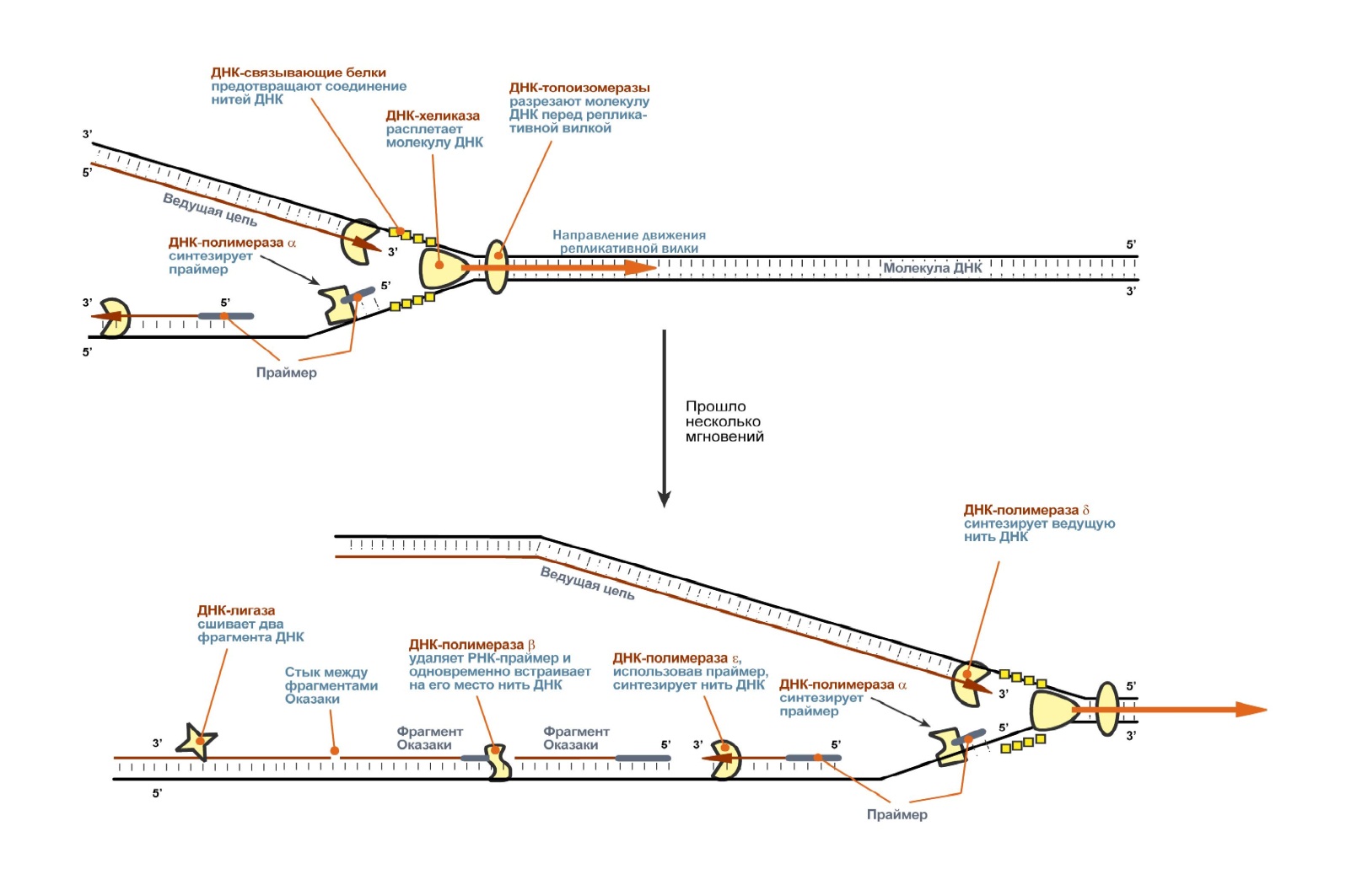
72. Репликация и репарация. В хронологическом порядке события репликации развертываются примерно следующим образом:
Повреждения и репарация ДНК Так как на геном любой неделящейся клетки постоянно оказывает влияние окружающая среда, то вполне вероятны повреждения в составе нуклеотида, также возможно встраивание неправильного нуклеотида при репликации. Такие нарушения быстро определяются специальными ферментами, пораженный участок удаляется, заполняется ДНК-полимеразой β и сшивается ДНК-лигазой. В случае изменения структуры основания (например, его дезаминирование) это основание удаляется ДНК-N-гликозидазой, затем другими ферментами удаляется дезоксирибоза и на ее место ДНК-полимеразой β и ДНК-лигазой встраивается нужный нуклеотид. Отрыв пуриновых и пиримидиновых оснований от дезоксирибозы устраняется ДНК-инсертазами, которые присоединяют к оставшейся дезоксирибозе соответствующие основания. 73. Строение РНК. Виды РНК. Их роль в метаболизме. Рибонуклеиновая кислота (РНК) – это однонитевой биополимер, в качестве мономеров которого выступают нуклеотиды. Матрицей для синтеза новых молекул РНК являются молекулы дезоксирибонуклеиновой кислоты (транскрипция РНК). Хотя в ряде случаев возможен и обратный процесс (образование новых ДНК на матрице РНК в ходе репликации некоторых вирусов). Также основой для биосинтеза РНК могут быть другие молекулы рибонуклеиновой кислоты (репликация РНК). В транскрипции РНК, происходящей в ядре клетки, участвует целый ряд ферментов, наиболее значимым из которых является РНК-полимераза. Структура РНК. Молекула имеет однонитевое строение. Полимер. В результате взаимодействия нуклеотидов друг с другом молекула РНК приобретает вторичную структуру, различной формы (спираль, глобула и т.д.). Мономером РНК является нуклеотид (молекула, в состав которой входит азотистое основание, остаток фосфорной кислоты и сахар (пептоза)). РНК напоминает по своему строению одну цепь ДНК. Нуклеотиды, входящие в состав РНК: гуанин, аденин, цитозин, урацил. Аденин и гуанин относятся к пуриновым основаниям, цитозин и урацил к пиримидиновым. В отличие от молекулы ДНК, в качестве углеводного компонента рибонуклеиновой кислоты выступает не дезоксирибоза, а рибоза. Вторым существенным отличием в химическом строении РНК от ДНК является отсутствие в молекуле рибонуклеиновой кислоты такого нуклеотида как тимин. В РНК он заменён на урацил. Функции РНК различаются в зависимости от вида рибонуклеиновый кислоты. 1) Информационная РНК (и-РНК). Иногда данный биополимер называют матричной РНК (м-РНК). Данный вид РНК располагается как в ядре, так и в цитоплазме клетки. Основное назначение – перенос информации о строении белка от дезоксирибонуклеиновой кислоты к рибосомам, где и происходит сбор белковой молекулы. Относительно небольшая популяция молекул РНК, составляющая менее 1% от всех молекул. 2) Рибосомная РНК (р-РНК). Самый распространенный вид РНК (около 90% от всех молекул данного вида в клетке). Р-РНК расположена в рибосомах и является матрицей для синтеза белковых молекул. Имеет наибольшие, по сравнению с другими видами РНК, размеры. Молекулярная масса может достигать 1,5 миллионов кДальтон и более. 3) Транспортная РНК (т-РНК). Расположена, преимущественно, в цитоплазме клетки. Основное назначение- осуществление транспорта (переноса) аминокислот к месту синтеза белка (в рибосомы). Транспортная РНК составляет до 10% от всех молекул РНК, располагающихся в клетке. Имеет наименьше, по сравнению с другими РНК- молекулами, размеры (до 100 нуклеотидов). 4) Минорные (малые) РНК. Это молекулы РНК, чаще всего с небольшой молекулярной массой, располагающиеся в различных участках клетки (мембране, цитоплазме, органеллах, ядре и т.д.). Их роль до конца не изучена. Доказано, что они могут помогать созреванию рибосомной РНК, участвуют в переносе белков через мембрану клетки, способствуют редупликации молекул ДНК и т.д. 5) Рибозимы. Недавно выявленный вид РНК, принимающие активное участие в ферментативных процессах клетки в качестве фермента (катализатора). 6) Вирусные РНК. Любой вирус может содержать только один вид нуклеиновой кислоты: либо ДНК либо РНК. Соответственно, вирусы, имеющие в своём составе молекулу РНК, получили название РНК-содержащие. При попадании в клетку вируса данного типа может происходить процесс обратной транскрипции (образование новых ДНК на базе РНК), и уже вновь образовавшаяся ДНК вируса встраивается в геном клетки и обеспечивает существование, а также размножение возбудителя. Вторым вариантом сценария является образование комплиментарной РНК на матрице поступившей вирусной РНК. В этом случае, образование новых вирусных белков, жизнедеятельность и размножение вируса происходит без участия дезоксирибонуклеиновой кислоты только на основании генетической информации, записанной на вирусной-РНК. 74. Биохимические механизмы синтеза РНК. Транскрипция (синтез РНК) Прежде чем начнут синтезироваться белки, информацию об их строении необходимо "достать" из ДНК и доставить ее к месту синтеза белков. Этим занимаются информационные или матричные РНК. Одновременно клетке нужны транспортеры аминокислот – транспортные РНК и структурные компоненты органелл, синтезирующих белок, – рибосомальные РНК. Вся информация о строении транспортных и рибосомальных РНК также находится в ДНК. Поэтому существует процесс переписывания или транскрипции данных с ДНК на РНК (англ. Transcription – переписывание) – биосинтез РНК на матрице ДНК. Как в любом матричном биосинтезе в транскрипции выделяют 5 необходимых элементов:
Биосинтез РНК происходит в участке ДНК, который называется транскриптон, с одного края он ограничен промотором (начало), с другого – терминатором (конец). РНК-полимеразы эукариот имеют по две больших субъединицы и несколько малых субъединиц. Стадии транскрипции Выделяют три стадии транскрипции: инициация, элонгация и терминация. Инициация Промотор содержит стартовый сигнал транскрипции – ТАТА-бокс. Так называется определенная последовательность нуклеотидов ДНК, связывающая первый фактор инициации ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции (матричная нить ДНК). Так как промотор ассиметричен ("ТАТА"), то он связывает РНК-полимеразу только в одной ориентации, что определяет направление транскрипции от 5'-конца к 3'-концу (5'→3'). Для связывания РНК-полимеразы с промотором необходим еще один фактор инициации – σ-фактор (греч. σ – "сигма"), но сразу после синтеза затравочного фрагмента РНК (длиной 8-10 рибонуклеотидов) σ-фактор отрывается от фермента. Другие факторы инициации раскручивают спираль ДНК перед РНК полимеразой. Элонгация Белковые факторы элонгации обеспечивают продвижение РНК-полимеразы вдоль ДНК и расплетают молекулу ДНК на протяжении примерно 17 нуклеотидных пар. РНК-полимераза продвигается со скоростью 40-50 нуклеотидов в секунду в направлении 5'→3'. Фермент использует АТФ, ГТФ, ЦТФ, УТФ одновременно в качестве субстрата и в качестве источника энергии. Терминация РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью белкового фактора терминации, так называемого ρ-фактора (греч. ρ – "ро"), от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая является первичным транскриптом, предшественником мРНК или тРНК или рРНК. 75. Биохимические механизмы синтеза белка. Трансляция (синтез белка) Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК. После переноса информации с ДНК на матричную РНК начинается синтез белков. Каждая зрелая мРНК несет информацию только об одной полипептидной цепи. Если клетке необходимы другие белки, то необходимо транскрибировать мРНК с иных участков ДНК. Биосинтез белков или трансляция происходит на рибосомах, внутриклеточных белоксинтезирующих органеллах, и включает 5 ключевых элементов: матрица – матричная РНК, растущая цепь – полипептид, субстрат для синтеза – 20 протеиногенных аминокислот, рибосомальные белки, рРНК и белковые факторы. источник энергии – ГТФ, Выделяют три основных стадии трансляции: инициация, элонгация, терминация. Инициация Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина. В начале этой стадии формируются два тройных комплекса: первый комплекс – мРНК + малая субъединица + ИФ-3, второй комплекс – метионил-тРНК + ИФ-2 + ГТФ. После формирования тройные комплексы объединяются с большой субъединицей рибосомы. В этом процессе активно участвуют белковые факторы инициации, источником энергии служит ГТФ. После сборки комплекса инициирующая метионил-тРНК связывается с первым кодоном АУГ матричной РНК и располагается в П-центре (пептидильный центр) большой субъединицы. А-центр (аминоацильный центр) остается свободным, он будет задействован на стадии элонгации для связывания аминоацил-тРНК. 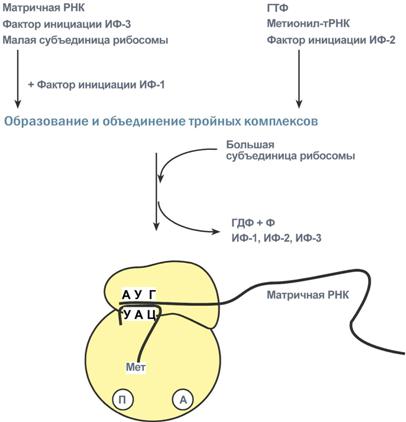 После присоединения большой субъединицы начинается стадия элонгации. Элонгация Для этой стадии необходимы все 20 аминокислот, тРНК для всех аминокислот, белковые факторы элонгации, ГТФ. Удлинение цепи происходит со скоростью примерно 20 аминокислот в секунду. Элонгация представляет собой циклический процесс. Первый цикл (и следующие циклы) элонгации включает три шага: Присоединение аминоацил-тРНК (еще второй) к кодону мРНК (еще второму),аминокислота при этом встраивается в А-центр рибосомы. Источником энергии служит ГТФ. Фермент пептидилтрансфераза осуществляет перенос метионина с метионил-тРНК (в П-центре) на вторую аминоацил-тРНК (в А-центре) с образованием пептидной связи между метионином и второй аминокислотой. При этом уже активированная СООН-группа метионина связывается со свободной NH2-группой второй аминокислоты. Здесь источником энергии служит макроэргическая связь между аминокислотой и тРНК. Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы, второй кодон (на рисунке) становится напротив П-центра, напротив А-центра оказывается третий кодон (на рисунке). Для этих процессов необходима затрата энергии ГТФ. Так как вместе с мРНК перемещаются закрепленные на ней тРНК, то инициирующая первая тРНК выходит из рибосомы, вторая тРНК с дипептидом помещается в П-центр. 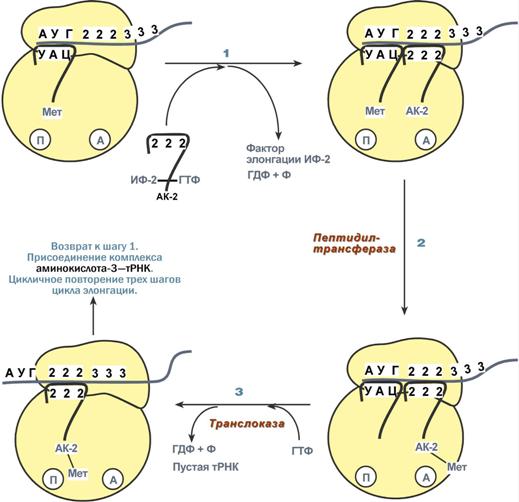 Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А-центр, после чего он смещается в П-центр в транслоказной реакции. В пустой А-центр входит четвертая аминоацил-тРНК и начинается третий цикл элонгации Образование пептидной связи при встраивании четвертой аминокислоты в пептид.Субъединицы рибосомы, большая часть транспортных РНК и матричная РНК не показаны. Цикл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь. Терминация Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют: Гидролитическое отщепление полипептида от конечной тРНК. Отделение от П-центра последней, уже пустой, тРНК. Диссоциацию рибосомы. Источником энергии для завершения трансляции является ГТФ. 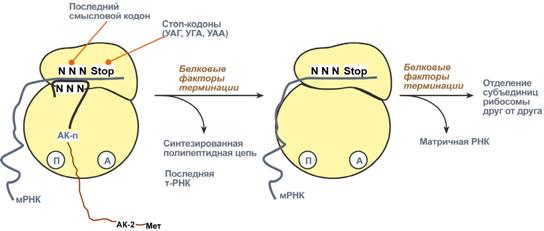 Реакции стадии терминации Полирибосомы По причине того, что продолжительность жизни матричной РНК невелика, перед клеткой стоит задача использовать ее максимально эффективно, т.е. получить максимальное количество "белковых копий". Для достижения этой цели на каждой мРНК может располагаться не одна, а несколько рибосом, встающих последовательно друг за другом и синтезирующих пептидные цепи. Такие образования называются полирибосомы. |