|
|
Экзамен по биохимии 2 курс. Вопросы итоговой аттестации по биохимии
Катаболические пути Пируват является продуктом окисления углеводов. Далее пируват превращается в ацетил-СоА и вовлекается в цикл трикарбоновых кислот. Кроме того, ацетил-СоА является также продуктом окисления жирных кислот, так что цикл трикарбоновых кислот участвует и в катаболизме жиров[24]. Стоит отметить, что пируват может вовлекаться в цикл трикарбоновых кислот и без превращения в ацетил-СоА, а преобразовавшись в малат под действием малат-фермента Анаболические пути Из α-кетоглутарата, образующегося в цикле трикарбоновых кислот, синтезируются аминокислоты глутамин, глутамат, пролин и аргинин. Сукцинил-СоА выступает в качестве предшественника при синтезе порфиринов и гема. Цитрат участвует в синтезе жирных кислот[en] и стеролов (из цитрата образуется ацетил-СоА, кроме того, он выступает в роли регулятора[26]). Малат может транспортироваться из митохондрий в цитоплазму, где он обратимо превращается в оксалоацетат. Получившийся оксалоацетат может служить предшественником для синтеза аминокислот аспартата, аспарагина, метионина, треонина и изолейцина, а также пиримидинов. Он может также превратиться в фосфоенолпируват с затратой GTP, а фосфоенолпируват (ФЕП) может служить предшественником в биосинтезе фенилаланина, тирозина,триптофана, серина, глицина и цистеина. Пируват, получающийся из ФЕП в процессе гликолиза, может стать предшественником аланина, лейцина и валина, а также может участвовать в глюконеогенезе[24][25]. Анаплеротические пути Промежуточные соединения цикла трикарбоновых кислот, покинувшие цикл и задействованные в синтезе различных соединений, замещаются за счёт специальныханаплеротических реакций[en]. В нормальных условиях реакции, в ходе которых промежуточные соединения цикла вовлекаются в другие метаболические пути, и реакции, возмещающие их уход, находятся в динамическом равновесии, поэтому концентрация промежуточных соединений цикла трикарбоновых кислот поддерживается постоянной
51. Биосинтез гликогена.
Тут проще будет изобразить эти реакции схемой.
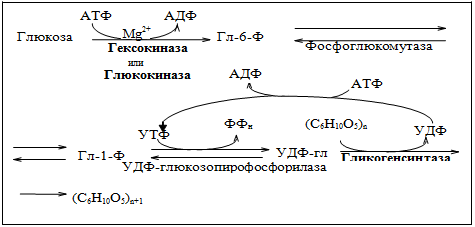
Первая реакция: Глюкоза под действием гексокиназы превращается в Глюкозо-6-фосфат
Вторая реакция: Глюкозо-6-фосфат превращается в Глюкозо-1-фосфат под действием фосфоглюкомутазы
В ходе третьей реакции Глюкозо-1-фосфат взаимодействуем с УТФ с образованием УДФ-глюкозы, фермент – УДФ-глюкозопирофосфорилаза.
И наконец, в ходе четвертой реакции из УДФ-глюкозы образуется гликоген под действием фермента Гликогенсинтазы.
52. Гликолиз и его биологическое значение.
Гликолиз – это цепь последовательных ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
Первой ферментативной реакцией гликолиза является фосфорилирование, т. е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой. В результате образуется глюкозо-6-фосфат.
Второй реакцией гликолиза является превращение глюкозо-6-фосфата под действием фермента гексозофосфатизомеразы во фруктозо-6-фосфат.
В третьей реакции образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ. Реакция катализируется ферментом фосфофруктокиназой. В результате образуется фруктозо-1,6-дифосфат.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-дифосфат расщепляется на две фосфотриозы - (дигидроксиацетонфосфата и глицеральдегид-3-фосфата.
Пятая реакция — реакция изомеризации триозофосфатов. Катализируется эта реакция ферментом триозофосфатизомеразой. В ходе этой реакции дигидроксиацентонфосфат превращается в глицеральдегид-3-фосфат.
В шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-дифосфоглицериновой кислоты и восстановленной формы НАД (НАДН2).
В седьмой реакции, которая катализируется фосфоглицераткиназой, происходит передача богатой энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты (3-фосфоглицерата).
В восьмой реакции происходит внутримолекулярный перенос оставшейся фосфатной группы и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
В девятой реакции 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват). Реакция катализируется ферментом енолазой.
В десятой реакции происходят разрыв высокоэргической связи и перенос фосфатного остатка от фосфоенолпировиноградной кислоты на АДФ, и образуется пировиноградная кислота. Катализируется эта реакция ферментом пируваткиназой.
В одиннадцатой реакции в результате восстановления пировиноградной кислоты образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН2+.
Биологическое значение процесса гликолиза прежде всего заключается в образовании богатых энергией фосфорных соединений. В первой стадии гликолиза затрачиваются две молекулы АТФ (гексокиназная и фосфофруктокиназная реакции). Во второй стадии образуются четыре молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции).
Таким образом, энергетическая эффективность гликолиза составляет две молекулы АТФ на одну молекулу глюкозы.
53. Глюконеогенез и его биологическая роль.
Глюконеогенез– синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пировиноградная кислоты, так называемые гликогенные аминокислоты, глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот.
Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза.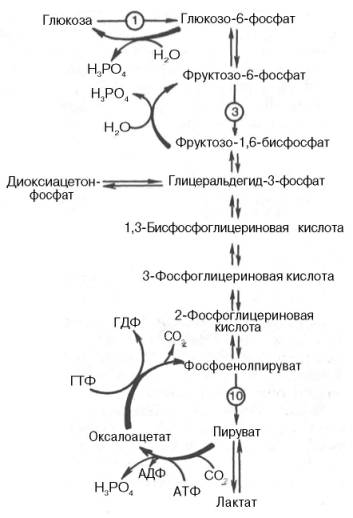
Образование из пирувата фосфоенолпирувата протеакает не напрямую, а через образование оксалоацетата.
Пируват → Оксалоацетат (фермент – пируваткарбоксилаза)
Оксалоацетат → Фосфоенолпируват (фермент – фосфоенопируваткарбоксилаза)
Затем идет обращение реакций гликолиза.
Реакция превращения Фруктозо-1,6-дифосфата в Фруктозо-6-фосфат происходит под действием фермента фруктозо-1,6-дифосфотазы.
И, наконец, реакция превращения Глюкозо-6-фосфата в глюкозу происходит под действием фермента глюкозо-6-фосфотазы.
(СХЕМА ЧИТАЕТСЯ СНИЗУ ВВЕРХ!!!)
54. Пентозофосфатный путь окисления углеводов.
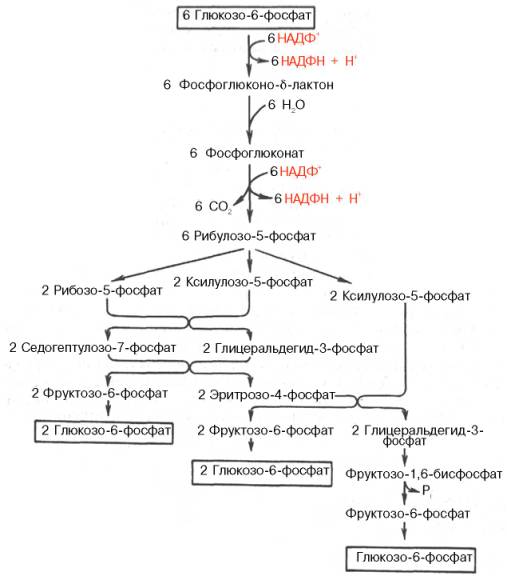
Пентозофосфатный цикл начинается с окисления глюкозо-6-фосфата и последующего окислительного декарбоксилирования продукта.
Это первая - окислительная стадия пентозофосфатного цикла.
Вторая стадия включает неокислительные превращения пентозофосфатов с образованием исходного глюкозо-6-фосфата.
Реакции пентозофосфатного цикла протекают в цитозоле клетки.
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ
Образовавшийся в ходе реакции 6-фосфоглюконо-δ-лактон – соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фосфоглюконовой кислоты (6-фосфоглюконат).
Во второй – окислительной – реакции, катализируемой 6-фосфоглюконатдегидрогеназой (декарбоксилирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН.
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза – ксилулозо-5-фосфат.
Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:
При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает так называемый неокислительный этап (стадия) пентозофосфатного цикла.
Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях.
В этих превращениях принимают участие ферменты: эпимераза, изомераза, транскетолаза и трансальдолаза
При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотриозы), а другие – специфические для пентозофосфатного пути (седогептулозо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов.
Коферментом в транскетолазной реакции служит ТПФ, играющий роль промежуточного переносчика гликольальдегидной группы от ксилулозо-5-фосфата к рибозо-5-фосфату. В результате образуется семиуглеродный моносахарид седогептулозо-7-фосфат и глицеральдегид-3-фосфат.
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз – при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритрозо-4-фосфатом.
Фермент трансальдолаза катализирует перенос остатка диоксиацетона от седогептулозо-7-фосфата на глицеральдегид-3-фосфат:
Эта реакция подобна реакции альдольного расщепления гликолитического пути, за исключением того, что в данном случае трёхуглеродный фрагмент, содержащий кетогруппу, переносится на альдосахар глицеральдегид-3-фосфат, а в гликолитическом пути кетофрагмент высвобождается в виде дигидроксиацетонфосфата.
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируется 5 молекул глюко-зо-6-фосфата.
55. Особенности углеводного обмена у жвачных животных. Пути синтеза глюкозы у жвачных животных.
У жвачных животных углеводный обмен имеет ряд особенностей. Углеводы у них всасываются из преджелудка преимущественно в виде летучих жирных кислот - уксусной, масляной и пропионовой. Уксусная кислота является предшественником молочного жира, используется организмом в качестве источника энергии и частично для синтеза глюкозы. Пропионовая кислота является основным источником глюкозы в организме жвачных. Использование масляной кислоты в обмене как источника энергии идет через стадию образования кетоновых тел. В норме в крови содержится 4- 6 мг%, в молоке -4-8, в моче - 9-10 мг% кетоновых тел. При заболевании коров кетозом уровень кетоновых тел повышается в крови до 19-50 мг%, в молоке - до 40, в моче - до 200- 300 мг%.
56. Роль летучих жирных кислот в метаболизме жвачных животных
Животные используют углеводы не только как источник энергии, но и в пластических целях для образования целого ряда структурных соединений: заменимых аминокислот, жирных кислот, гликопротеидов, гликолипидов, нуклеотидов и других. Углеводы составляют главную массу растительных кормов, которыми питаются травоядные животные. В сочных кормах углеводов содержится до 40%, в грубых - до 85%, в концентрированных - до 75%. Все углеводы, потребленные жвачными с кормом, в значительной части расщепляются или деструктируются в преджелудках микроорганизмами. Установлено, что в рубце подвергается разложению 60-65% клетчатки, 8095% легкопереваримых углеводов - крахмала и растворимых Сахаров
57. Строение клеточных мембран и их функции
Все живые клетки отделены от окружающей среды поверхностью называемой клеточной мембраной. Кроме того, для эукариотов характерно образование внутри клеток нескольких компартментов. Они представлены рядом субклеточных органелл, ограниченных мембранами, например, ядро и митохондрии. Мембраны представляют собой не только статически организованные поверхности раздела, но и включают активные биохимические системы, отвечающие за такие процессы, как избирательный транспорт веществ внутрь и наружу клетки, связывание гормонов и других регуляторных молекул, протекание ферментативных реакций, передача импульсов нервной системы и т.д. Существуют различные типы мембран, отличающиеся по выполняемым функциям.
Функции мембран: 1) граница 2) контроль транспорта метаболитов 3) рецепция сигналов и их передача 4) ферментные реакции 5) контакт с другими клетками
Химический состав: Мембраны состоят из липидных и белковых молекул, относительное количество которых варьирует у разных мембран. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5-10% вещества мембраны.
Липиды мембран:
Основная часть липидов в мембранах представлена фосфолипидами, гликолипидами и холестерином. Липиды мембран имеют в структуре две различные части: неполярный гидрофобный «хвост» и полярную гидрофильную «голову». Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных липидов, расположенных таким образом, что неполярные гидрофобные «хвосты» молекул находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются «хвост к хвосту» так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. Белки мембран включены в липидный двойной слой двумя способами: 1) связаны с гидрофильной поверхностью липидного бислоя - поверхностные мембранные белки 2) погружены в гидрофобную область бислоя - интегральные мембранные белки.
Транспорт веществ через мембраны: 1) Пассивный транспорт. 2) Простая диффузия 3) Облегченная диффузия. 4) Активный транспорт
58. Механизм транспорта липидов
Для преобразования энергии, заключенной в жирных кислотах в энергию существует метаболический путь окисления до CO2 и воды тесно связан и циклом Кребса. Этот путь называется бета-окислением, т.к. происходит окисление 3-го углеродного атома жирной кислоты. Эти реакции проходят в митохондриях большинства клеток организма (за исключением нервных). Для окисления используются кислоты, которые поступают в цитозоль из крови или появляющиеся при липолизе.
1 этап: Прежде чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активизироваться в цитозоле. Это происходит благодаря присоединению к ней коэнзима с образованием ацил-S-КоА, что является высокоэнергетическим соединением. Необратимость реакции происходит, когда образуется две молекулы фосфорой кислоты.
2 этап: Ацил- S- КоА не способен проходить через митохондриальную мембрану, поэтому перенос производится благодаря сцеплению с карнитином. После сцепления жирная кислота переносится транслоказой. Затем, на внутренней стороне мембраны ацил-S-КоА возвращается в исходную форму и начинается, собственно, бета – окисление.
3 этап: Процесс состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит 1) окисление (ацил-SКоА – дегидрогиназа), 2) гидратирование (еноил – SкоА- гидратаза), и вновь 3) окисление з-го атома углерода. В 4) реакции, трансферазной, от жирной кислоты ощепляется ацетил – SкоА.
59. β-Окисление жирных кислот
β-Окисление - специфический путь катаболизма жирных кислот, при котором от карбоксильного конца жирной кислоты последовательно отделяется по 2 атома углерода в виде ацетил-КоА. Метаболический путь - β-окисление - назван так потому, что реакции окисления жирной кислоты происходят у β-углеродного атома. Реакции β-окисления и последующего окисления ацетил-КоА в ЦТК служат одним из основных источников энергии для синтеза АТФ по механизму окислительного фосфорилирования. β-Окисление жирных кислот происходит только в аэробных условиях.
Активация жирных кислот
Перед тем, как вступить в различные реакции, жирные кислоты должны быть активированы, т.е. связаны макроэргической связью с коферментом А:
RCOOH + HSKoA + АТФ → RCO
КоА + АМФ + PPi.
Реакцию катализирует фермент ацил-КоА син-тетаза. Выделившийся в ходе реакции пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2 Н3РО4.
Выделение энергии при гидролизе макроэргической связи пирофосфата смещает равновесие реакции вправо и обеспечивает полноту протекания реакции активации.
Ацил-КоА синтетазы находятся как в цитозоле, так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к жирным кислотам с различной длиной углеводородной цепи. Жирные кислоты с короткой и средней длиной цепи (от 4 до 12 атомов углерода) могут проникать в матрикс митохондрий путём диффузии. Активация этих жирных кислот происходит в матриксе митохондрий. Жирные кислоты с длинной цепью, которые преобладают в организме человека (от 12 до 20 атомов углерода), активируются ацил-КоА синтетазами, расположенными на внешней мембране митохондрий.
β-Окисление жирных кислот - специфический путь катаболизма жирных кислот, протекающий в матриксе митохондрий только в аэробных условиях и заканчивающийся образованием ацетил-КоА. Водород из реакций β-окисления поступает в ЦПЭ, а ацетил-КоА окисляется в цитратном цикле, также поставляющем водород для ЦПЭ. Поэтому β-окисление жирных кислот - важнейший метаболический путь, обеспечивающий синтез АТФ в дыхательной цепи.
β-Окисление начинается с дегидрирования ацил-КоА FAD-зависимой ацил-КоА дегидрогеназой с образованием двойной связи между α- и β-атомами углерода в продукте реакции - еноил-КоА. Восстановленный в этой реакции кофермент FADH2 передаёт атомы водорода в ЦПЭ на кофермент Q. В результате синтезируются 2 молекулы АТФ (рис. 8-27). В следующей реакции р-окисления по месту двойной связи присоединяется молекула воды таким образом, что ОН-группа находится у β-углеродного атома ацила, образуя β-гидроксиацил-КоА. Затем β-гидроксиацил-КоА окисляется NАD+-зависимой дегидрогеназой. Восстановленный NADH, окисляясь в ЦПЭ, обеспечивает энергией синтез 3 молекул АТФ. Образовавшийся β-кетоацил-КоА подвергается тиолитическому расщеплению ферментом тиолазой, так как по месту разрыва связи С-С через атом серы присоединяется молекула кофермента А. В результате этой последовательности из 4 реакций от ацил-КоА отделяется двухуглеродный остаток - ацетил-КоА. Жирная кислота, укороченная на 2 атома углерода, опять проходит реакции дегидрирования, гидратации, дегидрирования, отщепления ацетил-КоА. Эту последовательность реакций обычно называют "циклом β-окисления", имея в виду, что одни и те же реакции повторяются с радикалом жирной кислоты до тех пор, пока вся кислота не превратится в ацетильные остатки.
Продуктами каждого цикла β-окисления являются FADH2, NADH и ацетил-КоА.Хотя реакции в каждом "цикле" одни и те же, остаток кислоты, который входит в каждый последующий цикл, короче на 2 углеродных атома. В последнем цикле окисляется жирная кислота из 4 атомов углерода, поэтому образуются 2 молекулы ацетил-КоА, а не 1, как в предыдущих.
|
|
|
 Скачать 1.2 Mb.
Скачать 1.2 Mb.