|
|
Экзамен по биохимии 2 курс. Вопросы итоговой аттестации по биохимии
50. Механизм реакций и биологическая роль цикла Кребса. Цикл Кребса протекает в митохондриях клеток, где он тесно связан с процессами биологического окисления и окислительного фосфорилирования. Поэтому хотя цикл может «крутиться» и в отсутствии кислорода, практически он зависит от его поступления в ткани. Остановка тканевого дыхания из-за отсутствия кислорода приводит к накоплению восстановленных НАД и ФАД и к остановке цикла. Цикл Кребса занимает центральное место в метаболизме клетки, так как в нем завершаются процессы полного окисления углеводов, жиров и белков. Так как в этом цикле используется ацетил-КоА образующийся как в процессе распада глюкозы, так и жирных кислот, то между двумя этими метаболитами наблюдается конкуренция за использование цикла Кребса. Цикл Кребса выполняет не только катаболитическую функцию, но и анаболическую, так как в нем образуется ряд продуктов необходимых для образования важных метаболитов. Например, 2-кетоглутарат и оксалоацетат используются для образования глутамата и аспартата, цитрат -для синтеза внемитохондрального ацетил-КоА, используемого для образования жирных кислот, сукцинил-КоА -для биосинтеза гема и др. Интенсивное использование метаболитов цикла Кребса для процессов биосинтеза может привести к их недостатку для выполнения энергетических функций. Поэтому существует ряд реакций, восполняющих эту убыль. К их числу относятся реакции образования оксалоацетата из пирувата или (особенно у беспозвоночных) из фосфоенолпирувата, образование оксалоацетата и 2-кетоглутарата из аспартата и глутамата и др. Особенностью цикла Кребса является то, что промежуточные вещества в нем не накапливаются и не расходуются, при отсутствии обмена с остальными метаболическими путями, происходит распад только ацетильных остатков. Если цикл Кребса рассматривать изолирование, то его можно рассматривать, как своеобразный ферментный комплекс катализирующий расщепление ацетил-КоА. Поэтому чтобы ускорить окисление ацетил-КоА нужно ускорить оборот цикла Кребса. Отсюда следует, что для регуляции цикла Кребса один или несколько ферментов должны существовать в двух формах - активной и малоактивной. В цикле Кребса известны 3 таких фермента: цитратсинтаза, НАД-изоцитратдегидрогеназа и 2-оксоглута-ратдегидрогеназа. Активность этих ферментов регулируется KoA-SH и соотношением АТФ, АДФ и фосфатом неорганическим. цикл Кребса– общий конечный путьокисленияацетильныхгрупп (в виде ацетил-КоА), в которые превращается в процессекатаболизма большая часть органическихмолекул, играющих роль «клеточноготоплива»:углеводов,жирных кислоти аминокислот. Образовавшийся в результате окислительногодекарбоксилированияпирувата вмитохондрияхацетил-КоА вступает в цикл Кребса. Данный цикл происходит в матриксемитохондрийи состоит из восьмипоследовательных реакций .Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образованиялимонной кислоты(цитрата). Затемлимонная кислота(шестиуглеродное соединение) путем рядадегидрирований(отнятиеводорода) и двухдекарбоксилирований(отщепление СО2) теряет два углеродныхатомаи снова вцикле Кребсапревращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла однамолекулаацетил-КоА сгорает до СО2и Н2О, амолекулаокса-лоацетата регенерируется. Рассмотрим все восемьпоследовательных реакций(этапов)цикла Кребса. 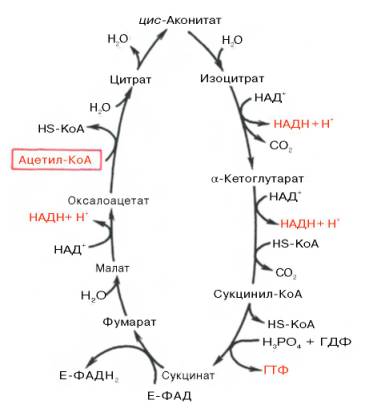 Первая реакциякатализируетсяферментомцит-рат-синтазой, при этомацетильнаягруппа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуетсялимонная кислота: По-видимому, в даннойреакциив качестве промежуточного продукта образуется связанный сферментомцитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованиемцитратаи HS-KoA. В результате второй реакцииобразовавшаясялимонная кислотаподвергается дегидратированию с образованием цис-аконитовойкислоты, которая, присоединяямолекулуводы, переходит визолимонную кислоту(изоцитрат). Катализирует эти обратимыереакциигидратации–дегидратацииферментаконитатгидратаза (аконитаза). В результате происходит взаимоперемещение Н и ОН вмолекулецитрата: Третья реакция, по-видимому, лимитирует скоростьцикла Кребса.Изолимонная кислотадегидрируется в присутствии НАД-зависимой изо-цитратдегидрогеназы. В ходе изоцитратдегидрогеназнойреакцииизолимонная кислотаодновременно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическимферментом, которому в качестве специфическогоактиваторанеобходимАДФ. Кроме того,ферментдля проявления своейактивностинуждается вионахMg2+или Мn2+. Во время четвертой реакциипроисходит окислительное декарбокси-лирование α-кетоглутаровойкислотыс образованием высокоэнергетического соединения сукцинил-КоА. Механизм этойреакциисходен с таковымреакцииокислительногодекарбоксилированияпирувата до ацетил-КоА, α-кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае вреакциипринимают участие 5коферментов: ТПФ, амидлипоевой кислоты, HS-KoA, ФАД и НАД+. Пятая реакциякатализируетсяферментомсукцинил-КоА-синтета-зой. В ходе этойреакциисукцинил-КоА при участии ГТФ инеорганического фосфатапревращается вянтарную кислоту(сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА: В результате шестой реакциисукцинатдегидрируется вфумаровую кислоту.Окислениесукцинатакатализируетсясукцинатдегидрогеназой, вмолекулекоторой сбелкомпрочно (ковалентно) связанкоферментФАД. В свою очередьсукцинатдегидрогеназапрочно связана с внутренней ми-тохондриальноймембраной: Седьмая реакцияосуществляется под влияниемферментафума-ратгидратазы (фумаразы). Образовавшаяся при этомфумаровая кислотагидратируется, продуктомреакцииявляетсяяблочная кислота(малат). Следует отметить, что фумаратгидратаза обладаетстереоспецифичностью(см. главу 4) – в ходереакцииобразуется L-яблочнаякислота: Наконец, в ходе восьмой реакциицикла трикарбоновых кислотпод влиянием митохондриальной НАД-зависимоймалатдегидрогеназыпроисходитокислениеL-малата в оксалоацетат. Восьмишаговый циклический процесс окисления простой ацетильной двухуглеродной группы до СO2 может показаться излишне сложным и не отвечающим биологическому принципу максимальной экономии. Однако роль цикла трикарбоновых кислот не исчерпывается окислением ацетат-иона (а значит, и углеводов, жирных кислот и некоторых аминокислот, при окислении которых он образуется). Этот путь составляет сердцевину метаболизма промежуточных соединений. Четырёх- и пятиуглеродные конечные продукты многих катаболических процессов вступают в цикл на промежуточных этапах. Оксалоацетат и α-кетоглутарат, например, являются продуктами катаболизма аспарагиновой и глутаминовой кислот, которые образуются при расщеплении белков. В некоторых метаболических процессах задействованы многие промежуточные соединения цикла, они служат предшественниками во многих анаболических процессах. Таким образом, цикл трикарбоновых кислот является амфиболическим путём, он связывает катаболические и анаболические процессы[23]. |
|
|
 Скачать 1.2 Mb.
Скачать 1.2 Mb.