№ 17 Первый и второй законы Менделя. Закон «чистоты гамет». Менделирующие признаки человека. Аутосомно - доминантный и аутосомно-рецесивный типы наследования.
Первый закон Менделя (единообразия гибридов) – при скрещивании гомозиготных родительских особей, которые отличаются по одной паре аллельных признаков, все гибриды первого поколения единообразны по фенотипу и генотипу.
Второй закон Менделя (расщепления гибридов второго поколения) – при моногибридном скрещивании гетерозиготных организмов у гибридов второго поколения происходит расщепление по фенотипу в отношении 3:1 и по генотипу – 1:2:1
Закон чистоты гамет - у гетерозиготной особи половые клетки «чисты», т.е. несут по одному гену из каждой аллельной пары.
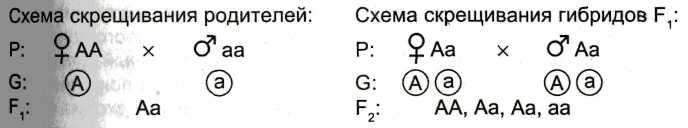
Аутосомно-доминантный тип наследования - классические примеры доминантного наследования – способность свертывать язык в трубочку и «свисающая» (свободная) мочка уха. Альтернативой последнему признаку является срощенная мочка – признак рецессивный. Еще одна наследственная аномалия у человека, обусловленная аутосомно-доминантным геном, – многопалость, или полидактилия. Она известна с глубокой древности. На картине Рафаэля «Сикстинская Мадонна» слева от Марии – папа римский Сикст II, на левой руке у него 5 пальцев, а на правой – 6. Отсюда и его имя: сикст – это шесть.
Аутосомно-рецессивный тип наследования - у человека описано очень много не сцепленных с полом признаков, которые наследуются как рецессивные. Например, голубой цвет глаз проявляется у людей, гомозиготных по соответствующему аллелю. Рождение голубоглазого ребенка у родителей с карими глазами повторяет ситуацию анализирующего скрещивания – в этом случае ясно, что они гетерозиготны, т.е. несут оба аллеля, из которых внешне проявляется только доминантный. Признак рыжих волос, определяющий еще и характер пигментации кожи, также является рецессивным по отношению к нерыжим волосам и проявляется только в гомозиготном состоянии.
|
№ 18 Третий закон Менделя. Цитологические основы универсальности законов Менделя. Менделирующие признаки человека.
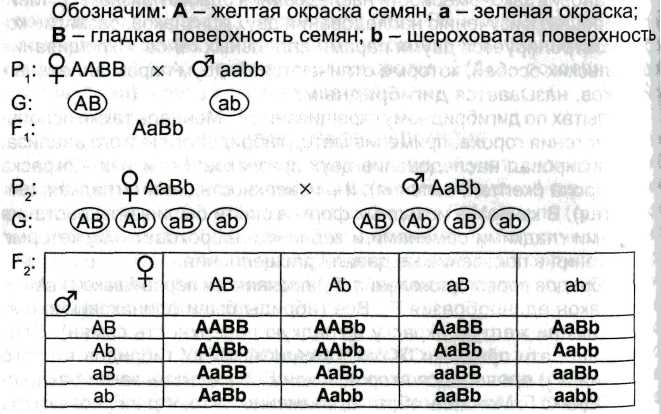
Третий закон Менделя (независимого наследования признаков) – при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
Закон проявляется, как правило, для тех пар признаков, гены которых находятся вне гомологичных хромосомах. Если обозначить буквой и число аллельных пар в негомологичных хромосомах, то число фенотипических классов будет определяться формулой 2n, а число генотипических классов - 3n. При неполном доминировании количество фенотипических и генотипических классов совпадает.
Цитологические основы законов Менделя базируются на:
1) парности хромосом (парности генов, обусловливающих возможность развития какого-либо признака)
2) особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися на них генами к разным пблюсам клетки, а затем и в разные гаметы)
3) особенностях процесса оплодотворения (случайного комбинирования хромосом, несущих по одному гену из каждой аллельной пары)
|
№ 19 Аллельные гены. Определение. Форма взаимодействия. Множественный аллелизм. Примеры. Механизм возникновения.
Аллельные гены - различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологических хромосом. Аллели определяют варианты развития одного и того же признака. В нормальной диплоидной клетке могут присутствовать не более двух аллелей одного локуса одновременно. В одной гамете два аллеля находиться не могут.
Форма взаимодействия аллельных генов.
Форма взаимодействия между аллельными генами может быть не только в виде доминирования (полное подавление проявления одного гена другим). Это наиболее простой способ наследования.
Взаимодействие генов с другими генами бывает таким:
• комплиментарное — совместное действие двух и более неаллельных генов (из разных пар) на проявление определенного признака. При этом каждый ген в отдельности не может вызвать развитие данного признака, только наличие в одном генотипе (гомо- или гетерозиготном состоянии) обуславливает его проявление;
• модифицирующее (гены-модификаторы) — усиление или ослабление действия главных генов действием других, неаллельных им генов (в некоторых породах ослабление черного окраса до голубого, коричневого до желтого);
• неаллельное — развитие признака под действием двух или более неаллельных генов;
• полимерное — суммарное выражение действия нескольких неаллельных генов на развитие одного и того же признака. Таким способом наследуется большинство количественных признаков (рост, вес, длина и др.);
• эпистатическое (эпистаз) — подавление доминантным аллелем одного гена действия другого, неаллельного первому гену (так ген Cd ослабляет красный цвет до желтого при наличии гена красного окраса);
• кодоминантность — независимое проявление признаков у гетерозиготы, контролируемых разными аллелями;
• плейотропия — способность одного гена влиять на проявление нескольких признаков (ген М, обеспечивающий мраморный окрас у догов, шелти, колли, такс, влияет также на цвет глаз, которые могут быть голубыми, голубо-карими, карими. При этом все варианты являются нормой).
Множественный аллелизм - наличие у гена множественных аллелей.
Создается так называемая серия аллелей, “рассеянных” в популяции данного вида. Итак, разнообразные стойкие состояния одного и того же гена, занимающего определенный локус в хромосоме, представленные то в виде нормального аллеля, то в виде мутации, получили название множественных аллелей. Примером множественного аллелизма может служит система групп крови АВО, открытая австрийским ученым К. Ландштейнером в 1900 г
|
№ 20 Наследование групп крови. Наследование резус-фактора. Резус конфликт.
Наследование групп крови.
В основе закономерностей наследования групп крови лежат следующие понятия. В локусе гена АВО возможны три варианта (аллеля) - 0, A и B, которые экспрессируются по аутосомно-кодоминантному типу. Это означает, что у лиц, унаследовавших гены А и В, экспрессируются продукты обоих этих генов, что приводит к образованию фенотипа АВ (IV). Фенотип А (II) может быть у человека, унаследовавшего от родителей или два гена А, или гены А и 0. Соответственно фенотип В (III) - при наследовании или двух генов В, или В и 0. Фенотип 0 (I) проявляется при наследовании двух генов 0. Таким образом, если оба родителя имеют II группу крови (генотипы AА или А0), кто-то из их детей может иметь первую группу (генотип 00). Если у одного из родителей группа крови A(II) с возможным генотипом АА и А0, а у другого B(III) с возможным генотипом BB или В0 - дети могут иметь группы крови 0(I), А(II), B(III) или АВ (IV).
Наследование резус-фактора.
Наследование резус-фактора кодируется тремя парами генов и происходит независимо от наследования группы крови. Наиболее значимый ген обозначается латинской буквой D. Он может быть доминантным - D, либо рецессивным - d. Генотип резус-положительного человека может быть гомозиготным - DD, либо гетерозиготным - Dd. Генотип резус-отрицательного человека может быть - dd.
Резус конфликт.
Гемолитическая болезнь плода и новорожденного это состояние, возникающее в результате несовместимости крови матери и плода по некоторым антигенам. Наиболее часто гемолитическая болезнь новорожденного развивается вследствие резус-конфликта. При этом у беременной женщины резус-отрицательная кровь, а у плода резус-положительная. Во время беременности резус-фактор с эритроцитами резус-положительного плода попадает в кровь резус-отрицательной матери и вызывает в ее крови образование антител к резус-фактору (безвредных для нее, но вызывающих разрушение эритроцитов плода). Распад эритроцитов приводит к повреждению печени, почек, головного мозга плода, развитию гемолитической болезни плода и новорожденного. В большинстве случаев заболевание быстро развивается после рождения, чему способствует поступление большого количества антител в кровь ребенка при нарушении целостности сосудов плаценты.
|
№ 21 Неаллельные гены. Формы их взаимодействия. Примеры.
Взаимодействия неаллельных генов.
1. Комплементарность. Этот вид взаимодействия генов заключается в том, что при наличии двух доминантных аллелей разных генов появляется новый признак, то есть для появления нового признака у организма должен быть генотип АВ. Так, для развития окраски необходимо, чтобы в организме синтезировались определенные белки и ферменты, превращающие их в пигмент. Классическим примером является наследование окраски цветков у душистого горошка.
2. Эпистаз. При эпистатическом взаимодействии одна пара генов может подавлять действие другой пары генов. Например, у лошадей масть определяется двумя парами генов. В одной паре генов доминантный аллель А определяет серую окраску (раннее поседение). Этот доминантный ген подавляет действие не только аллельного ему рецессивного гена а, но и подавляет проявление другой пары генов, определяющих масть (вороную, рыжую, гнедую), вне зависимости от того, является эта пара рецессивной или доминантной гомозиготой или гетерозиготой - окраска лошади будет только серой (лошади с генотипами ААвв, Аавв, ААВВ, АаВВ или АаВв).
3. Полимерия. Многие признаки определяются несколькими парами генов. Это характерно, в основном, для количественных признаков, таких как яйценоскость у кур, жирность молока у коров.
4. Плейотропное действие гена. При плейотропном действии гена один ген определяет развитие или влияет на проявление нескольких признаков. Это свойство генов было хорошо исследовано на мышах. Из схемы, видно, что ген определяет несколько признаков и признак определяется несколькими генами, поэтому можно сделать вывод, что плейотропное действие гена неразрывно связано с полимерным взаимодействием генов.
5. Летальные гены. Летальность генов - одна из разновидностей плейотропного действия гена. Так один ген, определяющий какой-либо признак, влияет так же на жизнеспособность в целом.
Ярким примером летальности гена служит ген платиновости у лисиц.
|
№ 22 Закон Моргана. Хромосомная теория наследственности.
Закон Моргана.
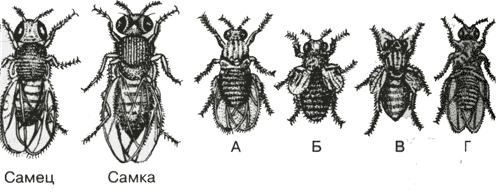
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обуславливающий развитие нормальных крыльев, - доминирует над геном недоразвыитых крыльев.
При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья - Сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т.е. они находятся в одной хромосоме. наследование сцепленных генов называют - сцепленное наследование.
Сцепление может нарушаться. Это доказывают особи В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.
Хромосомная теория наследственности.
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом;
6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
7. Каждый вид имеет характерный только для него набор хромосом - кариотип.
|
№ 23 Полное и неполное сцепление генов. Понятие о генетических картах хромосом. Метод соматической гибридизации клеток и его применение для картированных хромосом человека.
Полное и неполное сцепление генов.
Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть: полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна и неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Генетические карты хромосом.
Это схемы относительного расположения сцепленных между собой
наследственных факторов — генов. Г. к. х. отображают реально
существующий линейный порядок размещения генов в хромосомах (см. Цитологические карты хромосом) и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения.
Метод соматической гибридизации клеток.
При слиянии плазматических мембран клеток образуются клетки с двумя или большим числом ядер гетерокарионы. После первого деления ядра сливаются и образуется одно ядро с набором хромосом от всех слившихся партнеров образуется гибридная клетка. Низкую частоту образования гибридов можно было увеличить, использовав ряд агентов, вызывающих слияние: вирус Сендай, лизолецитин, полиэтиленгликоль.
Даже при использовании агентов, повышающих слияние, частота
образующихся гибридов крайне низка. Для их выделения необходимы селективные среды, позволяющие расти преимущественно образовавшимся гибридам. В настоящее время разработано несколько принципов селекции гибридных клеток. Одним из наиболее распространенных является метод, основанный на применении системы, содержащей гипоксантин, амидоптерин и тимидин (система ГАТ).
Так, с помощью методов соматической гибридизации клеток можно
определить участок хромосомы (обычно он располагается в области ее разрыва), в котором локализован тот или иной онкоген.
|
№ 24 Хромосомный механизм наследования пола. Цитогенетические методы определения пола. Наследование сцепленное с полом. Примеры.
Хромосомный механизм определения пола.
В клетках организмов содержится двойной набор гомологичных хромосом, которые называют аутосомами, и две половые хромосомы. В клетках женских особей содержатся две гомологичные половые хромосомы, которые принято обозначать XX. В клетках мужских особей половые хромосомы не являются парными – одна из них обозначается X, а другая Y. Таким образом, хромосомный набор у мужчин и женщин отличается одной хромосомой. У женщин в каждой клетке тела (кроме половых) 44 аутосомы и две половые хромосомы XX, а у мужчины – те же 44 аутосомы и две половые хромосомы Х и Y. При формировании половых клеток происходит мейоз и число хромосом в сперматозоидах и яйцеклетках уменьшается в два раза. У женщин все яйцеклетки имеют одинаковый набор хромосом: 22 аутосомы и X-хромосома. У мужчин образуются два вида сперматозоидов в соотношении 1:1 – 22 аутосомы и Х- или 22 аутосомы и Y-хромосома. Если при оплодотворении в яйцеклетку проникнет сперматозоид, содержащий Х-хромосому, появится зародыш женского пола, а если сперматозоид, содержащий Y-хромосому, – образуется зародыш мужского пола.
Таким образом, определение пола у человека, других млекопитающих, дрозофилл, зависит от наличия или отсутствия Y-хромосомы в сперматозоиде, оплодотворяющем яйцеклетку. Противоположная картина наблюдается у птиц и многих рыб: XY – набор половых хромосом самок, а XX – самцов. У некоторых насекомых, например, пчел, самки имеют XX-хромосомы, а у самцов только одна половая хромосома X, а парная ей отсутствует. Следовательно, в мире животных хромосомное определение пола может различаться.
Цитогенетический метод определения пола.
Цитогенетический метод основан на микроскопическом изучении хромосом в клетках человека. Сейчас используют метод дифференциального окрашивания хромосом, который позволяет точно идентифицировать хромосомы по характеру распределения в них окращшиваемых сегментов.
Наследование сцепленное с полом.
Половые хромосомы Х и Y содержат большое количество генов. Наследование определяемых ими признаков называют наследованием, сцепленным с полом, а локализацию генов в половых хромосомах называют сцеплением генов с полом.
Например, Х-хромосома человека содержит доминантный ген Н,
пределяющий свертывание крови. У человека, рецессивно гомозиготного по этому признаку, развивается тяжелое заболевание гемофилия, при котором кровь не сворачивается и человек может погибнуть от малейшего повреждения сосудов. Так как в клетках женщин две Х-хромосомы, то наличие в одной из них гена h не влечет за собой заболевания, так как во второй из них присутствует доминантный ген Н. В клетках мужчин есть только одна Х-хромосома. Если в ней присутствует ген h, то у мужчины разовьется гемофилия, так как Y-хромосома не гомологична Х-хромосоме и в ней нет гена Н или h.
|
№ 47 Наследственные болезни человека. Принципы лечения, методы диагностики и профилактики. Примеры.
Наследственные болезни.
Наследственные болезни могут быть вызваны нарушениями в отдельных генах, хромосомах или хромосомных наборах.
Хромосомные болезни возникают при изменении структуры хромосом:
- удвоении или выпадении участка хромосомы,
- повороте участка хромосомы на 180 0,
- перемещении участка хромосомы на негомологичную хромосому.
Впервые связь между аномальным набором хромосом и резкими отклонениями от нормального развития была обнаружена в случае синдрома Дауна.
Симптомы этого заболевания:
низкий рост, короткопалые, характерный разрез глаз, аномалии многих внутренних органов, специфическое выражение лица, умственная отсталость.
Индивидуальное течение наследственной болезни у каждого больного, даже при самой строгой оценке на идентичность мутаций как этиологического фактора, не вызываемой сомнений. Не бывает двух одинаковых больных, страдающих одним и тем же заболеванием. Для наследственных заболеваний в той же мере характерен клинической полиморфизм, как и для ненаследственных. Полиморфизм наследственных болезней выражается в разном времени появления симптомов или начала заболевания, разной степени выраженности болезненных проявлений, неодинаковых сроках летальных исходов. Вариации в проявлении наследственных болезней не ограничены только клиническими характеристиками. Они выражаются также в колебаниях значений биохимических, иммунологических и других показателей, которые входят в общее понятие фенотипа.
Генные болезни. Как видно из самого определения, этологическим фактором генных болезней являются генные мутации. У человека примерно 75 тысяч генов, и каждый ген вследствие мутации может обуславливать другое строение белка. Следовательно, количество наследственных болезней генной природы очень велико.
Хромосомные болезни. Все хромосомные болезни можно разделить на две большие группы: вызванные геномными мутациями, т.е. изменением числа хромосом (полиплоидии, анеуплоидии) при сохранении структуры последних, и обусловленные хромосомными мутациями, т.е. изменением структуры хромосомы (транслокации, делеции, инверсии).
Болезни с наследовательным предрасположением. Они отличаются от генных болезней тем, что для своего проявления нуждаются в действии факторов внешней среды и представляют собой наиболее обширную группу наследственной патологии, весьма многообразную по нозологическим формам. | |
 Скачать 284 Kb.
Скачать 284 Kb.