Микробиология, 1кт. микракт1. 5. Структурнофункциональная характеристика жгутиков, микроворсинок и включений. Методы их выявления
 Скачать 1.27 Mb. Скачать 1.27 Mb.
|

|
5. Структурно-функциональная характеристика жгутиков, микроворсинок и включений. Методы их выявления Жгутики бактерий определяют подвижность бактериальной клетки. Они представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12–20 нм, длина 3–15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка, базального тельца, содержащего стержень со специальными дисками 32 (1 пара дисков – у грамположительных и 2 пары – у грамотрицательных бактерий) и моторными белками (рис. 9.). 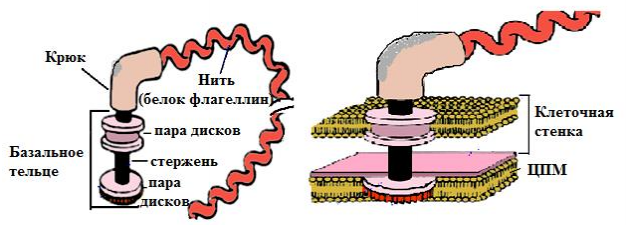 Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем – ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. У бактерий жгутики правовращающиеся. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер. Жгутики состоят из белка – флагеллина (от. лат. flagellum – жгутик), являющегося антигеном, – так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки (рис. 10). Подавляющее большинство наделенных жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчетов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны. У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счет броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения. 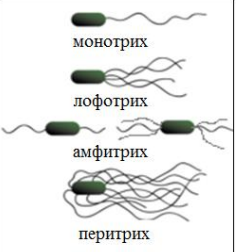 Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio. Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения). Ворсинки, или пили (фимбрии), – нитевидные образования, более тонкие и короткие, чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, который организован в спиральную структуру. Архитектура пилей варьирует от тонких нитевидных до толстых прочных палочковидных образований с осевыми отверстиями. Они обладают антигенной активностью. Различают пили общего типа (они обеспечивают прикрепление (адгезию) бактерии к различным субстратам, а также отвечают за питание, водносолевой обмен) и половые (F-пили), или конъюгационные, пили. Пили общего типа имееют более глубокую классификацию. Так, пили I типа располагаются перитрихиально, то есть по всей поверхности бактерии. У одной клетки может быть 50–400 пилей длиной до 1,5 мкм. Формирование пилей общего типа I определяется генами, расположенными в хромосоме. Их активность подвержена фазовым вариациям (ген может быть активен либо нет). В культуре присутствуют как клетки, имеющие много пилей общего типа I, так и лишенные их. Размножению клеток, лишенных пилей, способствует выращивание культуры на агаре, тогда как клетки с пилями получают преимущество при выращивании культуры в жидкой среде без аэрации. При этом они образуют пленку. Пили типа I придают бактериям гидрофобность, снижают их электрофоретическую подвижность. Они вызывают агглютинацию эритроцитов за счет того, что такие бактерии приклеиваются к эритроцитам (так же, как к другим клеткам животных), а также к клеткам растений и грибов, к неорганическим частицам. В присутствии маннозы нарушаются гемагглютинация и прикрепление бактерий к животным клеткам вообще, поскольку пили типа I прикрепляются к поверхностным рецепторам, содержащим маннозу. В присутствии маннозы соответствующие участки пилей заняты ее молекулами. Адгезивность пилей зависит также от гидрофобности образующего их белка пилина. С маннозными рецепторами реагируют участки пилей, расположенные по всей их поверхности, тогда как за гидрофобные взаимодействия ответственны окончания пилей (рис. 11). Пили II типа сходны с пилями I типа, но не вызывают агглютинации эритроцитов, не способствуют образованию бакРис. 10. Расположение жгутиков 34 териями пленки в жидкой среде. Антигенно они близки к пилям I типа и представляют собой их мутантную форму. Описан и еще ряд вариантов пилей, близких к пилям I типа. Связи пилей общего типа I с патогенностью у штаммов Е. coli не удается обнаружить. У энтеропатогенных штаммов обычно образуются другие пили, кодируемые плазмидными генами. Известно несколько типов таких пилей, причем обнаруживается связь типа пилей со специфичностью бактерий в отношении тех или иных животных. 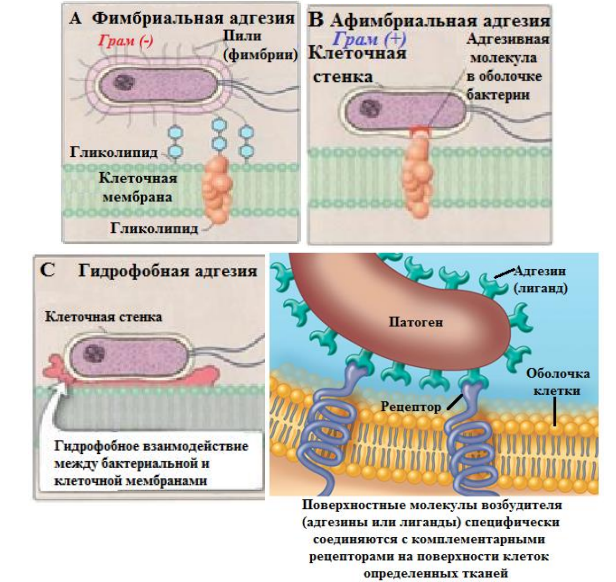 Экспрессия пилей вносит существенный вклад в колонизацию и персистенцию патогенных микроорганизмов в организме человека. Например, уропатогенные E. сoli для эффективного заселения эпителия мочевого пузыря должны иметь пили I типа, которые прикрепляются к консервативным рецепторам клеток эпителия мочевого пузыря, содержащим маннозу, и предотвращают вымывание бактерий с мочой. Р-пили выполняют ту же функцию в почках, препятствуя удалению пиелонефритных E. сoli из почек и мочевыводящих 35 каналов. Основное предназначение пилей – поддерживать на своих концах специфические прикрепительные структуры (адгезины). Адгезины являются посредниками при бактериальных контактах, при контактах с неживыми объектами, тканями и клетками восприимчивых организмов. Колонизация тканей хозяина бактериальными патогенами обычно зависит от стереохимического подобия между архитектурой адгезина и соответствующего рецептора клетки хозяина (см. рис. 11). Считают, что изучение функционирования пилей позволит не только глубже понять механизм колонизации и передачи сигналов, но и вести разработку новых поколений антимикробных препаратов. F-пили – половые пили, обычно 1–3 на клетку; они образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Соl-плазмиды). Отличительной особенностью половых пилей является их взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Все запасные вещества присутствуют в клетке в химически инертной форме. Такое состояние препятствует нарушению осмостаза клеточного содержимого. Некоторые включения просто лежат в цитоплазме, другие окружены тонкой мембраной. Мембрана обычно белковой природы, но иногда может содержать и липиды. Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в красно-фиолетовый цвет, а цитоплазма бактерии – в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронно-плотных гранул размером 0,1–1,0 мкм. 6. Особенности строения клеточной стенки бактерий. Клеточная стенка – важный структурный компонент бактериальной клетки. Клеточнаястенкавыполняетследующиефункции: определяет и сохраняет постоянную форму клетки; защищает внутреннюю часть клетки от действия механических и осмо- тических сил внешней среды; участвует в регуляции роста и деления клеток; обеспечивает коммуникации с внешней средой через каналы и поры; несет на себе специфические рецепторы для бактериофагов; определяет во многом антигенную характеристику бактерий; содержащийся в ее составе пептидогликан наделяет клетку важными иммунобиологическими свойствами; нарушение синтеза клеточной стенки бактерий является главной причи- ной их L-трансформации. Строениеклеточнойстенки.Основу клеточной стенки составляет пеп- тидогликан, который ранее называли муреином (от лат. murus – стенка). Он имеется только у эубактерий (кроме микоплазм). Пептидогликан включает остов и два набора пептидных цепочек – бо- ковых и поперечных (рис. 6). Остовпептидогликана одинаков у всех бактерий и состоит из чередующихся молекул аминосахаров – N-ацетилглюкозамина и N-ацетилмурамовой кислоты, связанных между собой β-гликозидными свя-зями. N-ацетилмурамовые кислоты связаны с тетрапептидами, которые сшиты между собой при помощи коротких пептидов (сшивка производится фермен- том транспептидазой). 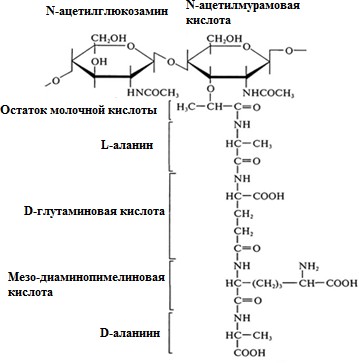 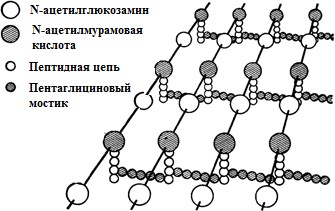 Рис. 6. Строение пептидогликана Боковыецепочкив каждой молекуле пептидогликана представлены набо- ром идентичных тетрапептидов. Поперечные цепочки также представлены набором из идентичных для данной молекулы пептидогликана пентапептидов, содержащих глицин (пентаглицины). У разных видов бактерий боковые и по- перечные пептиды различны. Пептидная цепочка содержит L-аланин, D-аланин, D-глутаминовую кис- лоту, L-лизин или мезо-диаминопимелиновую кислоту (только у грамотрица- тельных бактерий). Мезо-диаминопимелиновая кислота и D-аминокислоты в составе клеточных структур встречаются только в прокариотических клетках. Соседние тетрапептиды соединены между собой: у грамположительных мик- робов D-аланин связан через пентаглициновый мостик с L-лизином, у грамот- рицательных он соединяется непосредственно с мезо-диаминопимелиновой кислотой (рис. 7). 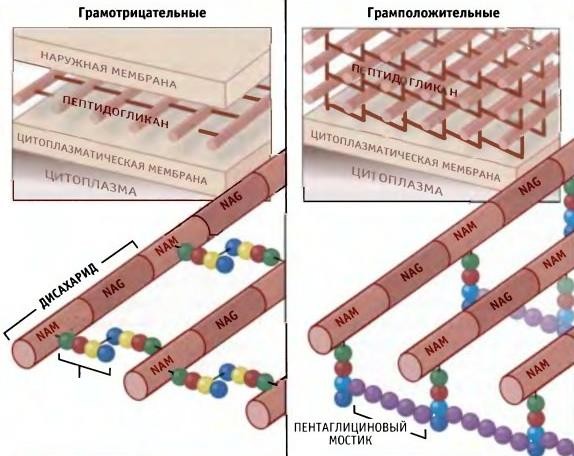 Рис. 7. Соединение тетрапептидов в молекулах пептидогликанов у грамотрицательных и грамположительных бактерий Наличие двух типов связей (гликозидные и пептидные), которые соеди- няют субъединицы пептидогликанов, придает этому гетерополимеру струк- туру молекулярной сети. Благодаря этим связямпептидогликановый слой кле- точной стенки образует огромного размера ригидную мешковидную макромо-лекулу, которая окружает протопласт, уравновешивает его тургорное давление и придает ему определенную постоянную форму. Пептидогликанможетраз-рушаться под действием различных ферментов, а его синтез блокируютбета-лактамные антибиотики. Это приводит к разрыхлению пептидоглика- новой сети, следствием чего является осмотический лизис растущих клеток. Пептидогликан, помимо того что он определяет постоянную форму бактерий, обладает следующими важнейшими иммунобиологическими свойствами. В его составе обнаружены родоспецифические антигенные детерми- нанты, локализованные в гликановом остове и в тетрапептидах. В межпептид- ных мостиках имеются видоспецифические антигенные детерминанты. Пептидогликан запускает классический и альтернативный пути актива- ции системы комплемента. Он тормозит фагоцитарную активность макрофагов и защищает бак- терии, особенно грамположительные, от фагоцитоза. Угнетает миграцию макрофагов. Способен индуцировать развитие гиперчувствительности замедлен- ного действия. Обладает противоопухолевым действием. Оказывает пирогенное действие на организм человека и животных. Состав и строение клеточной стенки – важный систематический признак, по которому все прокариоты подразделяются на следующие группы: толсто- стенные (Firmicutes – грамположительные – по отношению к окраске по ме- тоду Грама), тонкостенные (Gracilicutes – грамотрицательные) и без клеточ- ной стенки (Tenericutes) (рис. 8.) 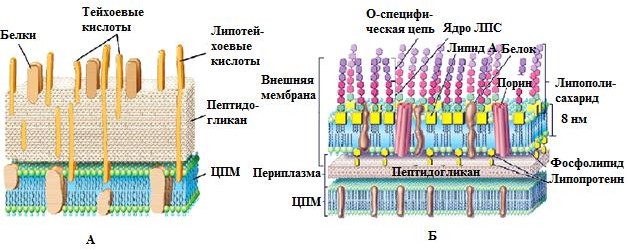 Рис. 8. Клеточная стенка грамположительных (А) и грамотрицательных (Б) бактерий |