1. Биология. Жизнь, происхождение. Уровни организации жизни. Термин биология
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Структура популяции – это определенная ее организация, формирующаяся на основе биологического свойства вида и под влиянием абиотических факторов среды и популяций других видов (межвидовая конкуренция). Структура популяции не стабильна. Различают следующие структуры популяции: пространственная; половая; генетическая; возрастная; поведенческая (этологическая). Половая структура популяции представляет собой соотношение в ней особей разного пола. Возрастная структура популяции — соотношение в составе популяции особей разного возраста, представляющих один или разные приплоды одного или нескольких поколений. Генетическая структура популяции определяется изменчивостью и разнообразием генотипов, частотами вариаций отдельных генов — аллелей, а также разделением популяции на группы генетически близких особей, между которыми при скрещивании происходит постоянный обмен аллелями. Пространственная структура популяции - характер размещения и распределения отдельных членов популяции и их группировок в ареале. Пространственная структура популяций заметно различается у оседлых и кочующих или мигрирующих животных. Экологическая структура популяции представляет собой разделение всякой популяции на группы особей, по-разному взаимодействующие с факторами среды. Экологическая характеристика популяции: Размеры ареала зависят от радиусов индивидуальной активности организмов данного вида и особенностей природных условий на соответствующей территории. Численность особей в популяциях организмов разных видов различается. Возрастная структура популяций организмов разных видов варьирует в зависимости от продолжительности жизни, интенсивности размножения, возраста достижения половой зрелости. В зависимости от вида организмов она может быть то более, то менее сложной. Половой состав популяций обусловливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов. Генетические характеристики популяции: Генетически популяция характеризуется ее генофондом (аллелофондом). Он представлен совокупностью аллелей, образующих генотипы организмов данной популяции. Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Генетическое единство популяции обусловливается достаточным уровнем панмиксии. Закон Харди-Вайнберга: В пределах генофонда популяции доля генотипов, содержащих разные аллели одного гена; при соблюдении некоторых условий из поколения в поколение не изменяется. Эти условия описываются основным законом популяционной генетики, сформулированным в 1908 г. английским математиком Дж. Харди и немецким врачом-генетиком Г. Вайнбергом. «В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствие мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение». Соответственно особи данной популяции образуют р гамет с аллелем А1 и q гамет с аллелем А2. Если скрещивания происходят случайным образом, то доля половых клеток, соединяющихся с гаметами А1, равна р, а доля половых клеток, соединяющихся с гаметами A2, — q. Возникающее в результате описанного цикла размножения поколение F1 образовано генотипами AlA1, A1A2, A2A2, количество которых соотносится как (р + q) (р + q) = р2 + 2pq + q2 Так как частоты гамет с разными аллелями в поколении FI в сравнении с родительским поколением не изменены, поколение F2 будет представлено организмами с генотипами AlA1, A1A2 и А2А2 в том же соотношении р2 + 2pq + q2 . Закон Харди — Вайнберга описывает условия генетической стабильности популяции. В законе Харди — Вайнберга перечислены условия, закономерно изменяющие генофонды популяций. К указанному результату приводят, например, факторы, ограничивающие свободное скрещивание (панмиксию), такие, как конечная численность организмов в популяции, изоляционные барьеры, препятствующие случайному подбору брачных пар. Генетическая инертность преодолевается также благодаря мутациям, притоку в популяцию или оттоку из нее особей с определенными генотипами, отбору. Человеческая популяция - группа людей, занимающих общую территорию и свободно вступающих в брак. В формировании популяций людей главную роль играют родственные связи. Демографические показатели (размер, уровень рождаемости и смертности, возрастной состав, экон.состояние, уклад жизни) оказывают серьезное воздействие на состояние генофондов человеческих популяций через структуру браков. Мутационный процесс у человека сходен с таковым у других организмов. В настоящее время давления мутационного процесса на генофонд человечества усиливается благодаря росту индуцированных мутаций (производственная деятельность, НТР = ионизирующее излучение). Доза в 1 Гр (грей) при низком уровене радиации: муж. 1000-2000 мутаций на миллион живых новорожденных, жен = 900 мутаций; следовательно повышение частоты злокачественных опухолей (лейкозы, рак молочной железы, щитовидной железы). Популяционные волны. Численность населения возросла с 5 миллионов в эпоху неолита до 6 миллиардов, но изменялась неравномерно. Ускорение прироста - развитие земледелия, индустриализации, НТР, изменения плотности населения (1чел на 3 км2 – 4-900-1200 на 3 км2). 50% населения размещаются на 5% площади суши - неравномерное распределения людей. Ускорение роста численности при ограниченности заселяемой территории способствует усилению миграции. Причина снижения - эпидемия чумы, опасные инфекции. Естественный отбор в человеческих популяциях утратил функцию видообразования, т.к биологические факторы исторического развития сменились социальными. Выполняет функцию стабилизации генофонда и поддержания наследственного разнообразия. 63. Естественный отбор, его формы. Роль естественного отбора в возникновении адаптаций и видообразовании. Особенности естественного отбора в человеческих популяциях. Естественный отбор — процесс, посредством которого в популяции увеличивается число особей, обладающих максимальной приспособленностью (наиболее благоприятными признаками), в то время как количество особей с неблагоприятными признаками уменьшается. В свете современной синтетической теории эволюции естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов. Естественный отбор — единственная известная причина адаптаций, но не единственная причина эволюции. К числу неадаптивных причин относятся генетический дрейф, поток генов и мутации. Термин «Естественный отбор» популяризовал Чарльз Дарвин, сравнивая данный процесс с искусственным отбором, современной формой которого является селекция. Идея сравнения искусственного и естественного отбора состоит в том, что в природе так же происходит отбор наиболее «удачных», «лучших» организмов, но в роли «оценщика» полезности свойств в данном случае выступает не человек, а среда обитания. К тому же, материалом как для естественного, так и для искусственного отбора являются небольшие наследственные изменения, которые накапливаются из поколения в поколение. Формы Е.О Движущий отбор — форма естественного отбора, которая действует при направленном изменении условий внешней среды. Описали Дарвин и Уоллес. В этом случае особи с признаками, которые отклоняются в определённую сторону от среднего значения, получают преимущества. При этом иные вариации признака (его отклонения в противоположную сторону от среднего значения) подвергаются отрицательному отбору. В результате в популяции из поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию). Примером действия движущего отбора является «индустриальный меланизм» у насекомых. «Индустриальный меланизм» представляет собой резкое повышение доли меланистических (имеющих тёмную окраску) особей в тех популяциях насекомых (например, бабочек), которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц, а тёмные — хуже. В XX веке в ряде районов доля тёмноокрашенных бабочек в некоторых хорошо изученных популяциях березовой пяденицы в Англии достигла 95 %, в то время как впервые тёмная бабочка (morfa carbonaria) была отловлена в 1848 году. Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определённом направлении, перемещая соответственно и норму реакции. Например, при освоении почвы как среды обитания у различных неродственных групп животных конечности превратились в роющие. Стабилизирующий отбор — форма естественного отбора, при которой его действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака. Понятие стабилизирующего отбора ввел в науку и проанализировал И. И. Шмальгаузен.Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детёнышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью. Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорождённые с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорождённые со средним весом. Учёт размера крыльев у воробьёв, погибших после бури в 50-х годах под Ленинградом, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи. Дизруптивный (разрывающий) отбор — форма естественного отбора, при которой условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дарвин описывал действие дизруптивного отбора, считая, что он лежит в основе дивергенции, хотя и не мог привести доказательств его существования в природе. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования.Одна из возможных в природе ситуаций, в которой вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам. Примером дизруптивного отбора является образование двух рас у погремка большого на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая. Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись только особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора. Половой отбор — это естественный отбор на успех в размножении. Выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Дарвин назвал это явление половым отбором. «Эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола». Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения, значительно выше, чем их недостатки для выживания. Положительный и отрицательный отбор Существует две формы естественного отбора: Положительный и Отсекающий (отрицательный) отбор. Положительный отбор увеличивает в популяции число особей, обладающих полезными признаками, повышающими жизнеспособность вида в целом. Отсекающий отбор выбраковывает из популяции подавляющее большинство особей, несущих признаки, резко снижающие жизнеспособность при данных условиях среды. С помощью отсекающего отбора из популяции удаляются сильно вредные аллели. Также отсекающему отбору могут подвергаться особи с хромосомными перестройками и набором хромосом, резко нарушающими нормальную работу генетического аппарата. Роль Е.О в возникновении адаптаций и видообразовании В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленностьопределяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора .Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции. Е.О. в человеческих популяциях Необходимая предпосылка отбора - борьба за существование: конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность.В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов.Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов. Естественный отбор в популяции людей действует так же, как и у других организмов, но имеет особенности: - действует слабее из-за развития медицины - действует в эмбриональном периоде или в раннем постэмбриональном периоде - потерял видообразовательную ф-ю (преобладает стабилизирующий отбор) - делает генофонд людей стабильным и разнообразным. Отбор против гетерозигот: примером является резус-антиген. Около 80% людей имеют в эритроцитах антиген D. Они резус-положительные Rh+. За синтез антигена отвечает доминантный аллель D.Резус-положительные люди имеют генотип DD или Dd. Резус-отрицательные люди имеют генотип dd. Если резус-отрицательная женщина беременна резус-положительным ребенком, и во время родов эритроциты ребенка попадают в кровь к матери, то у нее в ответ на резус-фактор (антиген) вырабатываются антитела. При второй беременности резус-положительным ребенком, антитела проникают через плаценту в организм ребенка и разрушают его эритроциты. Ребенок может умереть. Т.к. генотип ребенка Dd, отбор направлен против гетерозигот.Отбор против гомозигот рассмотрим на примере серповидноклеточной анемии. Есть аллель отвечающий за синтез нормального гемоглобина (HbA), а есть аллель отвечающий за синтез измененного гемоглобина (HbS). Изменённый гемоглобин обладает интересным свойством: в эритроцитах с таким гемоглобином плохо размножается малярийный плазмодий (люди с таким генотипом в 13 раз реже болеют малярией). 64. Дрейф генов как фактор эволюции, его действие в популяциях людей. Дрейф генов – случайное ненаправленное изменение частот аллелей в популяции. Обычно это явление характерно для малых популяций, здесь действие случайных процессов приводит к замтеным последствиям. В малых популяциях частота мутантного аллеля меняется быстро, случайным образом, и с высокой вероятностью происходит либо утрата аллеля, либо его высокая концентрация. Пример: в искусственно созданных популяциях концентрация нормального (А) и мутантного (а) аллелей оставила 50%. Спустя несколько поколений оказалось, что в некоторых популяциях все особи стали гомозиготными по мутантному аллелю(а), в других популяциях он был вовсе утрачен, и, наконец, часть популяций содержала как нормальный, так и мутантный аллель. То есть в результате дрейфа генов: во –первых, может возрастать генетическая однородность популяции(гомозиготность), во-вторых, в популяции может утрачиваться аллель, снижающий жизнеспособность особей, в – третьих, может возрастать концентрация редких аллелей. Ещё один пример данного явления, связанный с популяцией людей. Случайный рост частоты одной какой-либо мутации обычно обусловливается преимущественным размножением в изолированных популяциях. Это явление называется «эффектом родоначальника». Он возникает, когда несколько семей создают новую популяцию на новой территории. В ней поддерживается высокая степень брачной изоляции, что способствует закреплению одних аллелей и элиминацию других. Последствия «эффекта» - неравномерное распределение наследственных заболеваний человеческих популяций на земле. 65.Генетический полиморфизм и генетический груз естественных и человеческих попу-ляций. Механизмы возникновения и поддержания генетического полиморфизма. Наличие в популяции нескольких равновесно сосуществующих генотипов в концентрации, превышающей по наиболее редкой форме 1%1, называют полиморфизмом. Наследственный полиморфизм создается мутациями и комбинативной изменчивостью. Он поддерживается естественным отбором и бывает адаптационным (переходным) и гетерозиготным (балансированным). Адаптационный полиморфизм возникает, если в различных, но закономерно изменяющихся условиях жизни отбор благоприятствует разным генотипам. Так, в популяциях двухточечных божьих коровок Adalia bipunctata при уходе на зимовку преобладают черные жуки, а весной—красные (рис. 11.7). Это происходит потому, что красные формы лучше переносят холод, а черные интенсивнее размножаются в летний период.Балансированный полиморфизм возникает, если отбор благоприятствует гетерозиготам в сравнении с рецессивными и доминантными гомозиготами. Так, в опытной численно равновесной популяции плодовых мух Drosophila melanogaster, содержащей поначалу много мутантов с более темными телами (рецессивная мутация ebony), концентрация последних быстро падала, пока не стабилизировалась на уровне 10% (рис. 11.8). Анализ показал, что в созданных условиях гомозиготы по мутации ebony и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи. Это и создает состояние устойчивого полиморфизма по соответствующему локусу. Балансированный полиморфизм придает популяции ряд ценных свойств, что определяет его биологическое значение. Генетически разнородная популяция осваивает более широкий спектр условий жизни, используя среду обитания более полно. В ее генофонде накапливается больший объем резервной наследственной изменчивости. В результате она приобретает эволюционную гибкость и может, изменяясь в том или ином направлении, компенсировать колебания среды в ходе исторического развития. В генетически полиморфной популяции из поколения в поколение рождаются организмы генотипов, приспособленность которых неодинакова. В каждый момент времени жизнеспособность такой популяции ниже уровня, который был бы достигнут при наличии в ней лишь наиболее «удачных» генотипов. Величину, на которую приспособленность реальной популяции отличается от приспособленности идеальной популяции из «лучших» генотипов, возможных при данном генофонде, называют генетическим грузом. Он является своеобразной платой за экологическую и эволюционную гибкость. Генетический груз — неизбежное следствие генетического полиморфизма. 66.Общий план строения Хордовых. Узловые моменты в прогрессивной эволюции хордовых и их рекапитуляции в онтогенезе человека. Хордовые представляют собой наиболее высокоорганизованный тип в животном мире. Их общими чертами являются следующие. 1. Внутренний осевой скелет представлен хордой, которая присутствует в эмбриогенезе у всех представителей типа, а у высших дополняется, а затем и замещается позвоночником. 2. Над хордой располагается центральная нервная система в виде нервной трубки с полостью — невроцелем. 3. В боковых стенках глотки находятся жаберные щели, соединяющие ее полость с внешней средой. У рыб и некоторых земноводных они сохраняются в течение всей жизни, у высших хордовых — только в эмбриональном периоде. 4. Тело построено метамерно. У низших хордовых и у зародышей высших сегментация распростраянется на все системы органов, у высших ярко выражена только в эмбриональном периоде. Позже частично сохраняется только в опорно- двигательном аппарате, нервной и кровеносной системах. 5. Органами поддержания равновесия и движения являются конечности, причем у низших хордовых большее значение имеют непарные, а у высших —парные6. 6.Общий план строения хордовых представлен на рис. 13.14. На спинной стороне расположена нервная трубка, под ней — хорда или заменяющий ее позвоночник. Глубже находится пищеварительная трубка с развивающейся из нее дыхательной системой, а под ней — вентральный пульсирующий кровеносный сосуд или сердце. По бокам от нервной трубки и хорды лежат сомиты, а по бокам от кишки — спланхнотомы, внутри которых расположен целом. 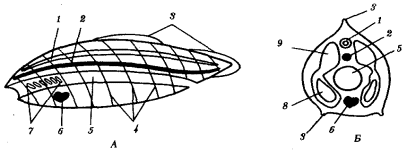 Рис. 13.14. Общий план строения хордового животного. А — вид сбоку; Б — поперечный срез: 1—нервная трубка, 2—хорда, 3—плавники, 4—сегменты тела, 5—пищеварительная трубка, 6—пульсирующий сосуд или сердце. 7—жаберные щели в глотке, 8—целом, 9—сомит На каких бы стадиях ни происходили эволюционные изменения, в онтогенезе обычно наблюдается известное повторение (рекапитуляция) развития предков. Это результат филогенетической обусловленности индивидуального развития.Онтогенез – не только предпосылка филогенетического развития, но и его результат. По этой причине изучение путей эволюционного процесса у современных животных и растений возможно путем анализа особенностей их эмбрионального развития.Изменения группы в филогенезе могут возникнуть лишь посредством изменений в онтогенезе. Обычно эти изменения индивидуального развития касаются поздних стадий развития. Самые же ранние стадии сохраняют значительное сходство с соответствующими ста-диями развития предковых и родственных форм – гласит Закон зародышевого сходства 67. Соотношение онто- и филогенеза. Закон зародышевого сходства К. Бэра. Биогенетический закон Ф. Мюллера - Э. Геккеля. Учение А.Н. Северцева о филэмбриогенезах. Индивидуальное развитие (онтогенез) – это совокупность процессов развития организма с момента образования зиготы и до смерти на основе реализации генетической информации в определенных условиях среды. Филогенез – это историческое развитие вида или другой биологической системы. К. Бэр в 1828 г. сформулировал 3 закона связи онто- и филогенеза: 1) закон зародышевого сходства: на ранних этапах эмбрионального развития зародыши различных животных в пределах типа сходны между собой (например, разных классов подтипа позвоночных). 2) закон последовательности появления признаков различного систематического ранга: первоначально в эмбриональном развитии появляются признаки типа, затем подтипа, класса, подкласса, отряда, семейства, рода, вида и, наконец, – индивидуальные. 3) закон эмбриональной дивергенции (расхождения признаков у зародышей): в процессе эмбриогенеза, по мере приобретения признаков различного систематического ранга, у зародышей различных животных в пределах типа возникают различия. В 1866г. Э.Геккель сформулировал биогенетический закон онтогенез– есть краткое и быстрое повторение филогенеза, обусловленное свойствами наследственности и приспособляемости. Закон вошел в биологию как закон Геккеля-Мюллера, так как Мюллер раньше дал формулировку закона, однако очень сложную. Мюллер также как Геккель обратил внимание, что в процессе развития могут быть рекапитуляции, а могут отсутствовать. Геккель в связи с разными типами развития выделил тип с палингенезами и ценогенезами. Палингенезы – это повторение у зародышей признаков их предков по филогенезу (закладка жаберных щелей, хорды, первичного хрящевого черепа, двухкамерного сердца у наземных позвоночных). Ценогенезы (эмбриоадаптации) – приспособительные признаки, которые возникают у зародышей и не сохраняются у взрослых организмов. Они нарушают ход исторического развития, по ним нельзя восстановить картину развития вида (развитие в эмбриогенезе у высших позвоночных амниона, хориона, аллантоиса). Дальнейшие эмбриологические исследования (А. Н. Северцов, И. И. Шмальгаузен) показали, что биогенетический закон верен лишь в общих чертах: 1 – нет ни одной стадии в развитии, на которой зародыш полностью повторял бы строение какого-либо предка по филогенезу; 2 – в онтогенезе повторяется строение не взрослых стадий предков, а их эмбрионов. Например, зародыш млекопитающих никогда целиком не повторяет строение рыбы, но на определенной стадии развития у него закладываются жаберные щели и жаберные артерии. Основное значение для объяснения связи онто- и филогенеза имеет учение А. Н. Северцова о филэмбриогенезах. Он рассматривал явление рекапитуляции с точки зрения эволюции онтогенезов. Филэмбриогенезы – это эмбриональные перестройки, которые сохраняются у взрослых форм и имеют адаптивное значение. А. Н. Северцов выделил 3 типа филэмбриогенезов: 1) архаллаксисы – это изменения с момента закладки органа (например, развитие волосяного покрова у млекопитающих); при этом в начале морфогенеза включаются мутировавшие гены и поэтому развитие идет новым путем (рекапитуляции отсутствуют); 2) девиации – уклонения с середины развития органа (например, развитие чешуи рептилий); первоначально повторяется формообразовательный процесс, характерный для предков по филогенезу, а в середине морфогенеза включаются в работу мутировавшие гены, и развитие органа идет новым путем (наблюдается частичная рекапитуляция); 3) анаболии – надставки, дополнения в развитии органа (например, от двухкамерного к четырехкамерному сердцу); первоначально рекапитулируют все предыдущие стадии развития органа, и только в конце эмбриогенеза включаются в работу мутировавшие гены, дополняющие формообразовательный процесс дальнейшей дифференцировкой (проявляется биогенетический закон). Эволюция чаще идет путем подбора анаболий, поэтому наблюдаются рекапитуляции, сходство эмбрионов на ранних стадиях развития и усложнение организации по мере появления новых систематических групп (типа хордовых, подтипа позвоночных, высших позвоночных и, наконец, человека). 68. Организм как целое в индивидуальном и историческом развитии. Способы преобразования органов в филогенезе Устойчивые взаимозависимости органов и систем, проявляющиеся в филогенезе, называют координациями. Механизмы соотносительных преобразований биологических структур различны, в связи с чем выделяют три их группы: биологические, динамические и топографические. Биологические координации наблюдаются между структурами, непосредственно не связанными ни по функциям, ни по месту положения. Основным связующим звеном между ними являются адаптации к определенным условиям обитания. Так, у большинства эндопаразитов сильно развиты половая система и органы прикрепления к телу хозяина, но при этом недоразвиты органы чувств и опорно-двигательный аппарат. Динамические координации выражаются во взаимном соответствии структур, связанных функционально. Тесные динамические координации имеются, например, между органами кровеносной и дыхательной систем. Так, животные, дышащие легкими, имеют трех- или четырехкамерное сердце и два круга кровообращения. Степень развитости нервных центров всегда соответствует интенсивности функционирования иннервируемых органов. Топографические координации проявляются между структурами, связанными друг с другом пространственно. Так, для каждого типа животного царства характерен своеобразный общий план строения, выражающийся в определенном взаимном расположении основных органов и систем. Например, у всех представителей типа Хордовые на спинной стороне тела расположена нервная трубка, под ней лежат хорда, пищеварительная трубка и брюшной кровеносный сосуд, а по бокам тела — производные мезодермы. Все типы координации характеризуются высокой степенью устойчивости. Так, хордовые животные, известные с конца протерозойской эры, сохранили неизменными общий план строения до настоящего времени, на протяжении более 500 млн. лет. Феномен паразитизма возник значительно раньше появления хордовых, и поэтому комплекс адаптации к паразитическому образу жизни является еще более древним. Высокая устойчивость филогенетических координации обеспечивается целостностью онтогенеза каждой конкретной особи, развитие всех биологических структур которой протекает в строгом взаимном соответствии. Такое соответствие структур развивающегося организма в онтогенезе называют онтогенетическими корреляциями. Различают геномные, эргонтические и морфогенетические корреляции. Геномные корреляции обеспечиваются целостностью генетической конституции развивающегося организма. Ведущими механизмами геномных корреляций являются генный баланс генотипа, сцепленное наследование генов, различные формы взаимодействия генов, а также плейотропность. Так, генные системы, регулирующие процессы пролиферации и избирательной гибели клеток на различных этапах органогенеза ,приводят к аллометрическому росту органов, благодаря чему появляются, например, удлиненный клюв, шея и задние конечности у большинства болотных птиц, длинная шея и ноги у жирафа, а также отличающиеся друг от друга пропорции тела у мужчин и женщин. Морфогенетические корреляции возникают между органами, пространственно связанными между собой. Они основаны либо на феномене эмбриональной индукции, либо на общности эмбриональных закладок органов. Эти корреляции проявляются уже на ранних стадиях онтогенеза, когда еще отсутствуют функциональные связи между формирующимися органами. Так, зачаток хорды обусловливает развитие нервной трубки на спинной стороне зародыша и дифференцировку скелетогенной ткани внутренних частей сомита — склеротома в хрящ или кость, а глазной бокал (вырост переднего мозга) — формирование хрусталика при морфогенезе глаза. Примером развития ряда структур из одного общего зачатка является формирование у млекопитающих и человека из закладок 1-й и 2-й пар жаберных дуг первичных челюстей, подъязычной кости, части хрящей гортани, шиловидного отростка черепа и трех слуховых косточек. Эргонтические корреляции обусловлены функциональной взаимозависимостью органов и частей организма. Они возникают на более поздних стадиях развития, когда органы начинают функционировать, и проявляются, например, в соответствии между степенью развитости мышцы, костных выступов, к которым она прикрепляется, и интенсивностью ее кровоснабжения. К такого рода корреляциям относят также соответствие вторичных половых признаков развитию гонад. Ведущие корреляции в онтогенезе — геномные. В конечном счете именно они лежат в основе других корреляций, значение которых на протяжении онтогенеза меняется. Это связано с первичностью изменений генотипа в процессе филогенеза. Между филогенетическими координациями и онтогенетическими корреляциями имеется теснейшая связь. Очевидно, что корреляции существуют и воспроизводятся в поколениях благодаря тому, что на протяжении предшествующей эволюции органов они преобразовывались скоординированно. С другой стороны, филогенетические координации в последующей эволюции организмов будут реализовываться благодаря воспроизведению онтогенетических корреляций в ходе индивидуального развития конкретных особей. Таким образом, в виде соотношения корреляций и координации проявляется диалектическое единство онто- и филогенеза как целостного процесса исторического развития живого. Осознание целостности и взаимообусловленности индивидуального и исторического развития необходимо врачу в связи с тем, что соотносительное преобразование биологических структур лежит в основе нормального развития организма человека, а нарушение даже одного незначительного компонента развивающейся системы может повлечь за собой возникновение целого комплекса нарушений в других ее элементах. Кроме филогенетических координации, подкрепляемых в каждом поколении онтогенетическими корреляциями, целостность развивающегося организма отражают и такие соотносительные преобразования органов, как субституция и гетеробатмия. Субституция — это такое эволюционное преобразование, при котором один орган замещается другим, выполняющим обычно ту же функцию с большей интенсивностью. При этом наблюдается развитие этих органов в разных направлениях. Один обычно подвергается редукции, другой — эволюционирует прогрессивно. Так, хорда замещается позвоночником и превращается в рудиментарное образование, а первичные хрящевые челюсти позвоночных заменяются вторичными костными. Это примеры гомотопной субституции, когда новый орган возникает на месте старого. При гетеротопной субституции заменяющий орган находится на новом месте. Так, функцию печени как органа кроветворения берет на себя красный костный мозг. Гетеробатмия — это такое эволюционное преобразование, при котором в одной группе организмов обнаруживается разный уровень эволюционной продвинутое и специализации разных частей одного и того органа, разных органов одной и той же системы или разных частей организма. Пример - человек, головной мозг которого за короткое время антропогенеза претерпел колоссальные изменения, в то время как пищеварительная система соответствует уровню развития других приматов. 69.Общие закономерности эволюционных преобразований органов. Гомологичные и аналогичные органы. Рудименты и атавизмы. В историческом развитии преобразования органов могут иметь прогрессивный или регрессивный характер. В первом случае органы увеличиваются в размерах и становятся более сложными по своему строению, во втором — уменьшаются в размерах, а их строение упрощается. Если у двух организмов, находящихся на разных уровнях организации, обнаруживаются органы, которые построены по единому плану, расположены в одинаковом месте и развиваются сходным образом из одинаковых эмбриональных зачатков, то это свидетельствует о родстве данных организмов. Такие органы называют гомологичными. Гомологичные органы часто выполняют одну и ту же функцию (например, сердце рыбы, земноводного, пресмыкающегося и млекопитающего), но в процессе эволюции функции могут и меняться (например, передних конечностей рыб и земноводных, пресмыкающихся и птиц). При обитании неродственных организмов в одинаковых средах у них могут возникать сходные приспособления, которые проявляются в возникновении аналогичных органов. Аналогичные органы выполняют одинаковые функции, строение же их, местоположение и развитие резко различны. Примерами таких органов являются крылья насекомых и птиц, конечности и челюстной аппарат членистоногих и позвоночных. Строение органов строго соответствует выполняемым ими функциям. При этом в исторических преобразованиях органов изменение функций непременно сопровождается и изменением морфологических характеристик органа. Основным принципом эволюции органических структур является принцип дифференциации. Дифференциация представляет собой разделение однородной структуры на обособленные части, которые в силу различного положения, связей с другими органами и различных функций приобретают специфическое строение. Таким образом, усложнение структуры всегда связано с усложнением функций и специализацией отдельных частей. Дифференцированная структура выполняет несколько функций, и строение ее сложно (Примером филогенетической дифференциации может являться эволюция кровеносной системы в типе хордовых). Отдельные части дифференцирующейся, ранее однородной структуры, специализируясь на выполнении одной функции, становятся функционально все более зависимыми от других частей данной структуры и от организма в целом. Такое функциональное соподчинение отдельных компонентов системы в целостном организме называют интеграцией (Четырехкамерное сердце млекопитающих представляет собой пример высокоинтегрированной структуры: каждый отдел выполняет лишь свою специальную функцию, не имеющую никакого смысла в отрыве от функций других отделов). |