1. Биология. Жизнь, происхождение. Уровни организации жизни. Термин биология
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Половое формирование проходит несколько этапов: Этап формирования генетического пола начинается в момент слияния яйцеклетки и сперматозоида. В зависимости от того, какую хромосому (Х или Y) несет сперматозоид, формируется женский или мужской генетический пол. Этап дифференцировки гонад определяет гонадный пол, и затем потенциальное направление репродуктивной функции индивидуума. Этап формирования гормонального пола определяется преобладанием тех или иных половых гормонов. В мужском организме количественно преобладают андрогены, а в женском – эстрогены. Этап формирования соматического пола заключается в развитии половых органов и вторичных половых признаков, характерных для данного биологического пола. Этап определения гражданского пола, то есть период рождения, когда, на основании внешних признаков новорожденного присваивают, тот или иной пол определяющий его полового воспитания. Классификация нарушений полового развития у человека: Нарушения хромосомного пола: Синдром Клайнфелтера; Мужчины с кариотипом ХХ; Дисгенезия гонад; Смешанная дисгенезия гонад (дефект развития половых желез); Истинный гермафродитизм. Нарушения гонадного пола: Чистая дисгенезия гонад; Синдром отсутствия тестикул. Нарушения фенотипического пола. Женский псевдогермафродитизм: Врожденная гиперплазия надпочечников; Женский псевдогермафродитизм вненадпочечникового генеза; Нарушения развития мюллеровых протоков. Мужской псевдогермафродитизм: Нарушения синтеза андрогенов; Нарушения действия андрогенов; Синдром персистенции мюллеровых протоков (одна из наиболее редких форм нарушения половой дифференцировки); Нарушение развития мужских гениталий. Морфологию хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы животных и растений в этом состоянии представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, у большей части хромосом удается легко найти зону первичной перетяжки, которая делит хромосому на два плеча. В области первичной перетяжки расположена центромера, или кинетохор. Это пластинчатая структура, имеющая форму диска. Она связана тонкими фибриллами с телом хромосомы в области перетяжки. Кинетохор является одним из центров полимеризации тубулинов, от него отрастают пучки микротрубочек митотического веретена, идущие в направлении к центриолям. Эти пучки микротрубочек принимают участие в движении хромосом к полюсам клетки при митозе. Некоторые хромосомы имеют вторичную перетяжку. Некоторые вторичные перетяжки представляют собой специализированные участки хромосом, связанные с образованием ядрышка. Плечи хромосом оканчиваются конечными участками – теломерами. Теломерные концы хромосом не способны соединяться с другими хромосомами или их фрагментами, в отличие от концов хромосом, лишенных теломерных участков (в результате разрывов), которые могут присоединяться к таким же разорванным концам других хромосом. По расположению первичной перетяжки (центромеры) выделяют следующие типы хромосом: 1. метацентрическая – центромера расположена посередине, плечи равной или почти равной длины, в метафазе приобретает V-образную форму; 2. субметацентрическая – первичная перетяжка слегка сдвинута к одному из полюсов, одно плечо немного длиннее другого, в метафазе имеет L-образную форму; 3. акроцентрическая – центромера сильно сдвинута к одному из полюсов, одно плечо гораздо длиннее другого, в метафазе не перегибается и имеет палочковидную форму; 4. телоцентрическая – центромера располагается на конце хромосомы, но такие хромосомы в природе не обнаружены. Обычно каждая хромосома имеет только одну центромеру (моноцентрические хромосомы), но могут встречаться хромосомы дицентрические (с 2-мя центромерами) и полицентрические (обладающие множеством центромер). Встречаются виды (например, осоки), у которых хромосомы не содержат видимых центромерных участков (хромосомы с диффузно расположенными центромерами). Они называются ацентрическими и не способны совершать упорядоченное движение при делении клетки. 42. Роль наследственности и среды в формировании пола организма. Пол организма представляет собой важную фенотипическую характеристику, которая проявляется в совокупности свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. Различают первичные и вторичные половые признаки. Под первичными половыми признаками понимают морфофизиологические особенности организма, обеспечивающие образование половых клеток — гамет, сближение и соединение их в процессе оплодотворения. Это наружные и внутренние органы размножения. Вторичными половыми признаками называют отличительные особенности того или другого пола, не связанные непосредственно с гаметогенезом, спариванием и оплодотворением, но играющие важную роль в половом размножении. Их развитие контролируется гормонами, синтезируемыми первичными половыми органами. Важным доказательством в пользу наследственной детерминированности половой принадлежности организмов является наблюдаемое у большинства видов соотношение по полу 1:1. Такое соотношение может быть обусловлено образованием двух видов гамет представителями одного пола (гетерогаметный пол) и одного вида гамет — особями другого пола (гомогаметный пол). Это соответствует различиям в кариотипах организмов разных полов одного и того же вида, проявляющимся в половых хромосомах. У гомогаметного пола, имеющего одинаковые половые хромосомы XX, все гаметы несут гаплоидный набор аутосом плюс Х-хромосому. У гетерогаметного пола в кариотипе кроме аутосом содержатся две разные или только одна половая хромосома (XY или ХО). Его представители образуют два вида гамет, различающиеся по гетерохромосомам: X и Y или X и 0. У большинства видов развитие признаков пола осуществляется на основе наследственной программы, заключенной в генотип. Однако известны примеры, когда половая принадлежность организма целиком зависит от условий, в которых он развивается (морской червь Bonellia viridis). У высших организмов значение среды в определении признаков пола, как правило, невелико. Обнаружение зависимости половой принадлежности развивающегося организма от дозы Х-хромосом у дрозофилы и некоторых других насекомых привело американского генетика С. Бриджеса (1922) к формулировке гипотезы генного баланса, в соответствии с которой организмы изначально бисексуальны. В процессе онтогенеза происходит выбор направления развития закладки в сторону признаков одного пола, включая дифференцировку половых желез, формирование половых путей и вторичных половых признаков. Первостепенная роль в развитии мужского или женского фенотипа принадлежит гормонам, образуемым гонадами. Генотип особи заключает в себе информацию о возможности формирования признаков того или иного пола, которая реализуется лишь при определенных условиях индивидуального развития. Изменение этих условий может стать причиной переопределения признаков пола. 43. Роль наследственности и среды в формировании пола организма. Для применения гибридологического метода генетического анализа, человек как вид обладает целым рядом особенностей, не позволяющих применять этот метод для изучения его наследственности и изменчивости. Во-первых, у человека не может быть произведено искусственного направленного скрещивания в интересах исследователя. Во-вторых, низкая плодовитость делает невозможным применение статистического подхода при оценке немногочисленного потомства одной пары родителей. В-третьих, редкая смена поколений, происходящая в среднем через 25 лет, при значительной продолжительности жизни дает возможность одному исследователю наблюдать не более 3—4 последовательных поколений. Изучение генетики человека затрудняется наличием в его геноме большого числа групп сцепления генов (23 у женщин и 24 у мужчин), а также высокой степенью фенотипического полиморфизма, связанного с влиянием среды. К методам, широко используемым при изучении генетики человека, относятся генеалогический, популяционно-статистический, близнецовый, метод дерматоглифики, цитогенетический, биохимический, методы генетики соматических клеток. Близнецовый метод. Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов. Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны, так как имеют 100% общих генов. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордантных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбрионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам между близнецами наблюдается дискордантность, т.е. различия. Напротив, сохранение сходства между близнецами, несмотря на различия условий их существования, свидетельствует о наследственной обусловленности признака. Применение: изучение наследственности и изменчивости у человека для определения соотносительной роли наследственности и среды в формировании различных признаков, как нормальных, так и патологических, оценить эффективность действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания). Популяционно-статистический метод С помощью популяционно-статистического метода изучают наследственные признаки в больших группах населения, в одном или нескольких поколениях. Существенным моментом при использовании этого метода является статистическая обработка получаемых данных. Этим методом можно рассчитать частоту встречаемости в популяции различных аллелей гена и разных генотипов по этим аллелям, выяснить распространение в ней различных наследственных признаков, в том числе заболеваний. Он позволяет изучать мутационный процесс, роль наследственности и среды в формировании фенотипического полиморфизма человека по нормальным признакам, а также в возникновении болезней, особенно с наследственной предрасположенностью. Этот метод используют и для выяснения значения генетических факторов в антропогенезе, в частности в расообразовании. При статистической обработке материала, получаемого при обследовании группы населения по интересующему исследователя признаку, основой для выяснения генетической структуры популяции является закон генетического равновесия Харди — Вайнберга. Он отражает закономерность, в соответствии с которой при определенных условиях соотношение аллелей генов и генотипов в генофонде популяции сохраняется неизменным в ряду поколений этой популяции. Математическим выражением закона Харди — Вайнберга служит формула (рА. + qa)2, где р и q — частоты встречаемости аллелей А и а соответствующего гена. Раскрытие этой формулы дает возможность рассчитать частоту встречаемости людей с разным генотипом и в первую очередь гетерозигот — носителей скрытого рецессивного аллеля: p2AA + 2pqAa + q2аа. 44. Особенности человека как объекта генетического анализа. Методы изучения генетики человека: генеалогический, биохимический, цитогенетический. Особенности человека как объекта генетических исследований Основные закономерности наследственности и изменчивости живых организмов были открыты благодаря разработке и применению гибридологического метода генетического анализа, основоположником которого является Г. Мендель. Наиболее удобными объектами, широко используемыми генетиками для гибридизации и последующего анализа потомства, стали горох, дрозофила, дрожжи, некоторые бактерии и другие виды, легко скрещивающиеся в искусственных условиях. Отличительной особенностью этих видов является достаточно высокая плодовитость, позволяющая применять статистический подход при анализе потомства. Короткий жизненный цикл и быстрая смена поколений позволяют исследователям в относительно небольшие промежутки времени наблюдать передачу признаков в последовательном ряду многих поколений. Немаловажной характеристикой видов, используемых в генетических экспериментах, является также небольшое число групп сцепления в их геномах и умеренное модифицирование признаков под влиянием окружающей среды. С точки зрения приведенных выше характеристик видов, удобных для применения гибридологического метода генетического анализа, человек как вид обладает целым рядом особенностей, не позволяющих применять этот метод для изучения его наследственности и изменчивости. Во-первых, у человека не может быть произведено искусственного направленного скрещивания в интересах исследователя. Во-вторых, низкая плодовитость делает невозможным применение статистического подхода при оценке немногочисленного потомства одной пары родителей. В-третьих, редкая смена поколений, происходящая в среднем через 25 лет, при значительной продолжительности жизни дает возможность одному исследователю наблюдать не более 3—4 последовательных поколений. Наконец, изучение генетики человека затрудняется наличием в его геноме большого числа групп сцепления генов (23 у женщин и 24 у мужчин), а также высокой степенью фенотипического полиморфизма, связанного с влиянием среды. Все перечисленные особенности человека делают невозможным применение для изучения его наследственности и изменчивости классического гибридологического метода генетического анализа, с помощью которого были открыты все основные закономерности наследования признаков и установлены законы наследственности. Однако генетиками разработаны приемы, позволяющие изучать наследование и изменчивость признаков у человека, несмотря на перечисленные выше ограничения. Невозможность направленного скрещивания, проводимого в интересах исследования, и малочисленность потомства, получаемого от каждой родительской пары, компенсируются подбором в популяции семей с интересующим генетика признаком в количестве, достаточном для проведения статистического анализа потомства. Ограниченность числа поколений, которые может наблюдать один генетик, компенсируется возможностью подбора и регистрации последовательных поколений семей с интересующим признаком многими поколениями исследователей. Существенно облегчается генетический анализ у человека благодаря высокой степени изученности его фенотипа методами морфологии, физиологии, биохимии, иммунологии, клиники. Большие перспективы в изучении наследственности и изменчивости у человека открываются в связи с применением ранее используемых и новых методов генетических исследований. Методы исследования генетики Биохимический метод; популяционно-статистический метод; молекулярно-генетические методы. Генеалогический метод основан на составлении родословной человека и изучении характера наследования признака. Суть его состоит в установлении родословных связей и определении доминантных и рецессивных признаков и характера их наследования. Особенно эффективен этот метод при исследовании генных мутаций. Этот метод используется для диагностики наследственных заболеваний и медико-генетического консультирования. По характеру наследования определяется вероятность рождения ребенка с генетическими аномалиями. Близнецовый метод основан на изучении фенотипа и генотипа близнецов для определения степени влияния среды на развитие различных признаков. Этот метод в 1876 г. предложил английский исследователь Ф. Гальтон для разграничения влияния наследственности и среды на развитие различных признаков у человека. Близнецовый метод позволяет определить степень проявления признака у пары, влияние наследственности и среды на развитие признаков. Все различия, которые проявляются у однояйцевых близнецов, имеющих одинаковый генотип, связаны с влиянием внешних условий. Таким образом, близнецовый метод позволяет выявить роль генотипа и факторов среды в формировании признака, для чего изучаются и сравниваются степени сходства (конкордантность) и различий (дискордантность) монозиготных и дизиготных близнецов. Цитогенетический метод заключается в микроскопическом исследовании структуры хромосом и их количества у здоровых и больных людей. Из трех типов мутаций под микроскопом могут обнаруживаться лишь хромосомные и геномные мутации. Биохимический метод основан на изучении характера биохимических реакций в организме, обмена веществ для установления носительства аномального гена или уточнения диагноза. Заболевания, в основе которых лежит нарушение обмена веществ, составляют значительную часть генной наследственной патологии. К ним относятся сахарный диабет, фенилкетонурия (нарушение обмена фенилаланина), галактоземия (нарушение усвоения молочного сахара) и другие. Этот метод позволяет установить болезнь на ранней стадии и лечить ее. 45. Медико-генетическое консультирование. Задачи, методы и этапы медико- генетического консультирования. Понятие о генетическом риске. Эффективных способов лечения наследственных заболеваний нет, поэтому основная роль отводится профилактике. Медико-генетическое консультирование – это один из видов специализированной помощи населению, направленный в первую очередь на предупреждение появления в семье детей с наследственной патологией. Консультирование (задачи) включает 3 этапа: 1. Уточнение диагноза (генетический анализ) 2. Прогноз потомства (теоретические расчеты, эмпирические данные). На этом этапе проводят проспективное (на ранних сроках) и ретропроспективное (после рождения). Генетический риск – это вероятность появления определенной наследственной патологии у обратившегося за консультацией или у его потомков. Риск до 5% - низкий, до 10% повышенный в легкой степени, средний – до 20%. Средний риск и выше – противопоказание к зачатию. 3. Врач объясняет степень риска и дает рекомендации. Показаниями для медико-генетического консультирования являются: 1) рождение ребенка с врожденным пороком развития; 2) установленная или подозреваемая наследственная болезнь в семье в широком смысле слова; 3) задержка физического развития или умственная отсталость у ребенка; 4) повторные спонтанные аборты, выкидыши, мертворождения; 5) близкородственные браки; 6) воздействие подозреваемых на тератогенность или известных тератогенов в первые 3 мес. беременности; 7) неблагополучное протекание беременности. 46. Онтогенез, определение и периодизация, типы онтогенеза. Основные периоды онтогенеза человека. Онтогене́з — индивидуальное развитие организма, совокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом, от оплодотворения (при половом размножении) или от момента отделения от материнской особи (при бесполом размножении) до конца жизни. Типы онтогенеза: Непрямое развитие – развитие с метаморфозом. Полный метаморфоз (личинка → куколка → особь) Неполный метаморфоз (личинка → особь) Прямое развитие: Неличиночное развитие (яйцекладущие – рептилии, земноводные, птицы) – из яйцевых оболочек развивается особь по строению такая же, как и взрослый организм. Внутриутробное развитие – формирование плода в метке женской особи (живородящие рептилии и млекопитающие). Периоды онтогенеза человека: Пренатальный – период внутриутробного развития Начальный период (консенсус): оплодотворение – до 3 суток и имплантация зиготы в эндометрий матки – ч/з 6-7 суток после оплодотворения. Зародышевый (эмбриональный период) – 8-9 недель Плодный период – от 9 недели – до начала схваток Постэмбриональный: Интранатальный – от схваток – первого вздоха Новорожденности – от первого вдоха, до 28 дней Грудной возраст – 28 дней – 12 месяцев Период молочных зубов: – Преддошкольный (ясельный) – 1-3 года – Дошкольный – 3-7 лет – Младший школьный – 7-12 лет – Старший школьный (подростковый или пубертатный период) – 12-14 – 16-18 лет Период половой зрелости – 16-18 – 40-45 лет Предменопаузы и менопауза – 40-45 – 50-55 лет Постменопауза Климактерический период Постэмбриональный период. Начинается с первого вдоха и началом функционирования легочного дыхания и малого круга кровообращения. Происходит адаптация к атмосферному воздуху, окружающей среде и ее флоре. В период новорожденности выявляются все пороки развития, которые устраняются хирургическим путем (порок сердца, расщелина губы, подвывих тазобедренного сустава), так де выявляются все мутации и последствия нарушения мейоза при гаметогенезе у родителей и митоза при делении зиготы. После созревания особи начинается гаметогенез Эмбриональный период. После попадания сперматозоидов в кислую среду влагалища, их большая часть гибнет, а оставшаяся часть проникает в матку, где щелочная среда способствует сохранению двигательной активности сперматозоидов. Через полчаса сперматозоиды достигают матки, через 1,5-2 часа они придвигаются к маточным трубам, где после оплодотворения яйцеклетки ее оболочка становится непроницаемой для других сперматозоидов, образуется зигота, которая по мере продвижения зиготы по маточной трубе к матке делится. 47.Строение и функции половых клеток. Морфофизиологические особенности яйцеклеток Хордовых, их типы. Связь строения яйца с типом дробления. Оплодотворение, его этапы. 1.Строение и функции половых клеток. Половые клетки имеют гаплоидный набор хромосом в ядрах, что обеспечивает воспроизведение в зиготе типичного для организмов данного вида диплоидного числа хромосом. Гаметы отличаются необычным для других клеток значением ядерно-цитоплазматического отношения. У яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша. У сперматозоидов благодаря малому количеству цитоплазмы ядерно-цитоплазматическое отношение высокое, т. к. главная задача мужской гаметы – транспортировка наследственного материала к яйцеклетке. Половые клетки отличаются низким уровнем обменных процессов, близким к состоянию анабиоза.По сравнению с другими клетками функция гамет уникальна. Они обеспечивают передачу наследственной информации между особями разных поколений, чем сохраняют жизнь во времени. 2.Морфофизиологические особенности яйцеклеток хордовых и их типы. Таблица 7.2. Типы яйцеклеток, встречающиеся у хордовых  Полилецитальные — содержат большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых).Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии).Олиголецитальные — содержат мало желтка (моллюски, иглокожие, млекопитающие. ).Алецитальные — не содержат желтка (некоторые паразитические перепончатокрылые).Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии).Гомо (изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные яйца (моллюски, иглокожие). Полилецитальные — содержат большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых).Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии).Олиголецитальные — содержат мало желтка (моллюски, иглокожие, млекопитающие. ).Алецитальные — не содержат желтка (некоторые паразитические перепончатокрылые).Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии).Гомо (изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные яйца (моллюски, иглокожие).4.Оплодотворение и его этапы Оплодотворение - это процесс слияния половых клеток, завершающийся объединением их генетического материала. Образующаяся в результате оплодотворения диплоидная клетка - зигота - представляет собой начальный этап развития нового организма. Процесс оплодотворения складывается из трех последовательных фаз: • дистантного взаимодействия и сближения гамет; • контактного взаимодействия гамет и активизации яйцеклетки; • слияния гамет, или сингамии. Первая фаза – сближение. Как при наружном (у рыб, амфибий), так и при внутреннем (у рептилий, птиц и млекопитающих) оплодотворении сперматозоиды в результате хемотаксиса в условиях слабощелочной среды очень быстро перемещаются по направлению к яйцеклеткам. Смещение рН в кислую сторону, наоборот, парализует спермии. Сперматозоиды млекопитающих обладают также реотаксисом, т.е. способностью двигаться против тока жидкости, направленного из яйцевода, где происходит оплодотворение, в матку. Кроме таксисов сближению половых клеток способствуют: перистальтика маточных труб и мерцательное движение ресничек эпителия маточных труб. Вторая фаза – проникновение сперматозоида через оболочки яйцеклетки. Контактное взаимодействие гамет наступает, когда сперматозоид сближается с яйцеклеткой. У млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Такое явление называется моноспермией. У беспозвоночных животных, рыб, амфибий, рептилий и птиц возможна полиспермия, когда в яйцеклетку проникает несколько сперматозоидов, но в слиянии ядер (оплодотворении) все равно принимает участие только один. Ферменты, выделяемые из акросом (трипсин, гиалуронидаза), разрушают лучистый венец, расщепляют гликозаминогликаны вторичной (блестящей) оболочки яйцеклетки. В цитоплазму яйцеклетки проникает головка, шейка и часть хвостового отдела (митохондриальное влагалище). Проникновение сперматозоида значительно усиливает процессы внутриклеточного обмена, что связано с повышением дыхания и активизацией ферментативных систем яйцеклетки. Третья фаза – образование мужского и женского пронуклеусов с последующим слиянием их (синкарион). При этом у многих видов животных ядра мужской и женской клеток во время сближения переходят в состояние метафазы. Затем хромосомы обоих ядер образуют единую материнскую «звезду», но уже с удвоенным (диплоидным) числом хромосом. В других случаях ядра вначале сливаются и затем переходят в состояние кариокинеза. Одновременно внесенные сперматозоидом центриоли расходятся к полюсам клетки, и этот одноклеточный зародыш – зигота вступает во второй период эмбрионального развития – период дробления. 3.Связь строения яйца с типом дробления 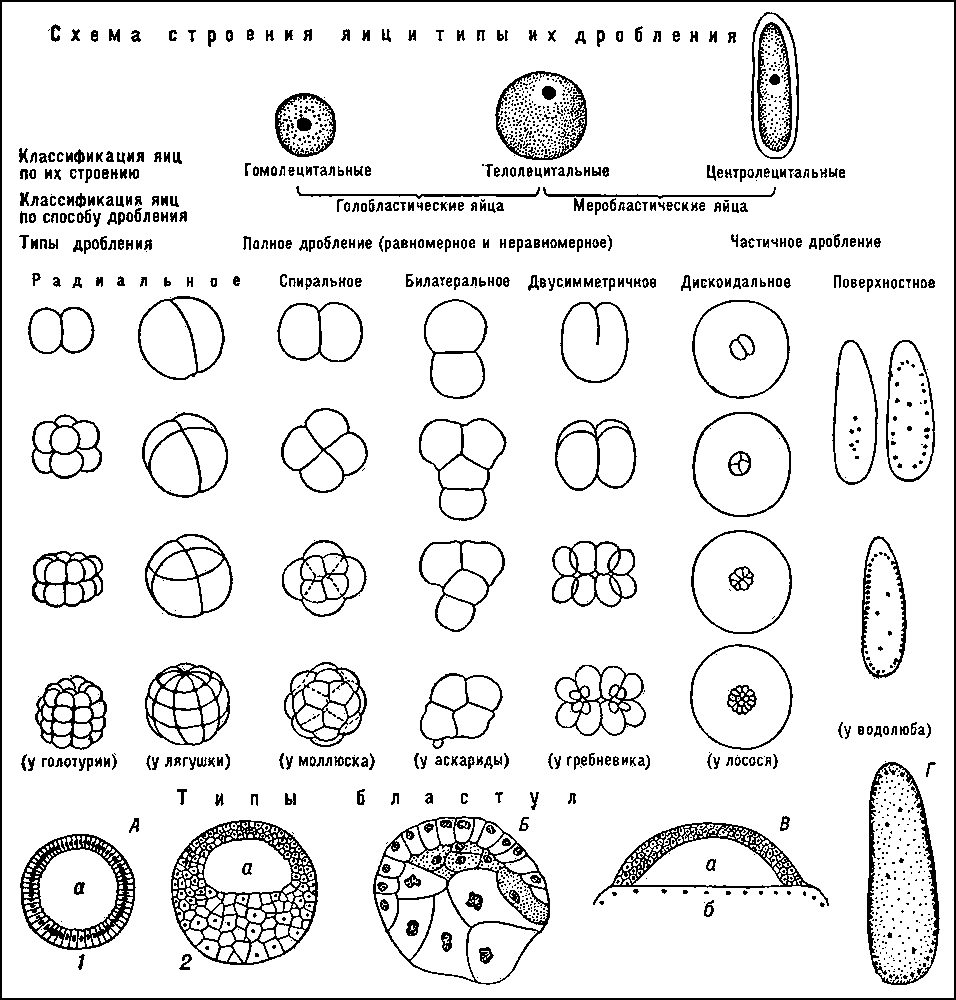 Схема строения яиц, типы их дробления и типы бластул: А — целобластула (1 — равномерная, 2 —неравномерная: а — бластоцель); Б — стерробластула; В — дискобластула (а — бластоцель, б — желток);Г — перибластула. Голобластическое дробление. Такое дробление характерно для яиц, содержащих сравнительно мало желтка, т. е. для олиго- и мезолецитальных, а также для умеренно телолецитальных яиц. Если в результате дробления образуются бластомеры примерно одинаковых размеров, говорят о равномерном дроблении, если же бластомеры явно различаются по величине — о неравномерном. Меробластическое дробление. Этот тип дробления наблюдается у животных с телолецитальными яйцами, которые отличаются высокой степенью концентрации желтка в вегетативной области. У головоногих моллюсков, многих рыб, а также у рептилий и птиц дробление происходит только в относительно небольшой части яйца, образующей как бы диск на поверхности яйцеклетки, — дискоидальное дробление. Абластическое дробление. Такое дробление характерно для центролецитальных яиц насекомых, поэтому его называют также централецитальным. В этом случае не происходит цитокинеза и деления цитоплазмы. Делятся только ядра, которые находятся в центральной области яйца, откуда они мигрируют по цитоплазматическим тяжам, пронизывающим яйцо, на поверхность. Радиальное дробление. У многих животных (книдарии, иглокожие, некоторые первично-хордовые, рыбы и амфибии) дробящееся яйцо имеет радиальную ось симметрии, при которой плоскость, проходящая через любой меридиан, делит зародыш на две геометрически тождественные половины. При радиальном дроблении два первых деления проходят во взаимно перпендикулярных меридиональных плоскостях, а третье — в экваториальной плоскости. Спиральное дробление. У аннелид и моллюсков в результате первых двух взаимно перпендикулярных меридиональных делений образуется стадия четырех бластомеров. Начиная с третьего деления дробления митотические веретена располагаются под некоторым углом к меридиональной плоскости. Благодаря этому образующиеся четыре клетки анимального полушария несколько смешаются относительно клеток вегетативного квартета и располагаются в промежутках между его бластомерами квадрантами. 48Клеточные механизмы онтогенеза. Классификация и механизмы формирования пороков развития у человека. Элементарные клеточные механизмы онтогенеза В онтогенезе особи происходят сложнейшие преобразования: осуществляется дифференциация частей развивающегося организма (морфогенез), формирование его внешней и внутренней структуры (морфогенез), рост. В основе этих преобразований лежат клеточные и системные механизмы развития. К клеточным механизмам относят размножение, перемещения, избирательную сортировку, дифференцировку, программированную гибель клеток. РАЗМНОЖЕНИЕ Деление клеток (размножение, пролиферация) играет важную роль в процессах онтогенеза. Во-первых, благодаря делению из зиготы, которая соответствует одноклеточной стадии развития, возникает многоклеточный организм. Во-вторых, пролиферация клеток, происходящая после стадии дробления, обеспечивает рост организма. В-третьих, избирательному размножению клеток принадлежит заметная роль в обеспечении морфогенетических процессов. В-четвертых, в постнатальном периоде индивидуального развития благодаря клеточному делению осуществляется обновление многих тканей в процессе жизнедеятельности организма (физиологическая или гомеостатическая регенерация), а также заживление ран, восстановление утраченных органов (репаративная регенерация) ПЕРЕМЕЩЕНИЯ В процессе развития особи происходят неоднократные перемещения (миграции) отдельных клеток, их групп, клеточных пластов. Особое значение миграция клеток приобретает на стадии гаструляции, приводя к формированию зародышевых листков. В ходе органогенеза этот механизм важен, например, при формировании крупных пищеварительных желез, производных нервного гребня. ИЗБИРАТЕЛЬНАЯ СОРТИРОВКА Механизм сортировки и слипания (адгезии) клеток лежит в основе выделения и объединения клеток одного типа среди всех прочих. В процессе развития клетки «узнают» друг друга и сортируются в зависимости от свойств, т.е. образуют скопления и пласты избирательно, только с определенными клетками. Этот механизм крайне важен при формировании зародышевых листков в ходе гаструляции, образовании структур в органогенезе, осуществлении регенеративных процессов и иммунных реакций в постнатальном развитии. Следует отметить, что перемещаться могут как отдельные клетки, так и целые клеточные пласты ДИФФЕРЕНЦИРОВКА Запрограммированный процесс реализации генетически обусловленной программы формирования специализированного фенотипа клеток, отражающего их способность к тем или иным профильным функциям. ГИБЕЛЬ процесс программированной гибели клеток - апоптоз. механизм программированной клеточной гибели обеспечивает регуляцию численности клеток, а именно - установление нужного равновесия между процессами пролиферации и гибели клеток, что в одних ситуациях обеспечивает стабильное состояние организма, в других - рост, в-третьих - атрофию тканей и органов. Пороки МЕХАНИЗМЫ РАЗВИТИЯ ЗАБОЛЕВАНИЯ Формирование пороков происходит преимущественно в период эмбрионального морфогенеза (3-10-я неделя беременности) в результате нарушения процессов размножения, миграции, дифференциации и гибели клеток. Эти процессы происходят на внутриклеточном, экстраклеточном, тканевом, межтканевом, органном и межорганном уровнях. Нарушением размножения клеток объясняют гипоплазию и аплазию органов. Нарушение их миграции лежит в основе гетеротопий. Задержка дифференциации клеток обусловливает незрелость или персистирование эмбриональных структур, а ее полная остановка - аплазию органа или его части. Нарушение физиологической гибели клеток, как и нарушение механизмов адгезии ("склеивание" и срастание эмбриональных структур), лежат в основе многих дизрафий (например, спинномозговых грыж). КЛАССИФИКАЦИЯ Выделяют несколько групп пороков. В зависимости от времени воздействия вредных факторов и объекта поражения выделяют следующие формы пороков развития. 1. Гаметопатии - патологические изменения в половых клетках, произошедшие до оплодотворения и приводящие к спонтанному прерыванию беременности, врожденным порокам развития, наследственным заболеваниям. 2. Бластопатии - это повреждения зиготы в первые 2 недели после оплодотворения вызывающие гибель зародыша, внематочную беременность, пороки развития с нарушением формирования оси зародыша 3. Эмбриопатии - поражения зародыша 15-й день после оплодотворения до 75-й день проявляющиеся пороками развития отдельных органов и систем, прерыванием беременности. Большинство врожденных пороков образуется именно в этот период. Локализация дефекта также зависит от интенсивности повреждающего воздействия. 4. Фетопатии - общее название болезней плода, возникающих под воздействием неблагоприятных факторов с 11-й недели внутриутробной жизни до начала родов. 49. Общая характеристика, сущность и основные клеточные механизмы дробления. Дробление у представителей Хордовых. Особенности дробления у плацентарных млекопитающих. Нарушения дробления. Экстракорпоральное оплодотворение у человека. Сущность дробления. Дробление- это ряд последовательных митотических делений зиготы и далее бластомеров, заканчивающихся образованием многоклеточного зародыша — бластулы. Возникающие при дроблении клетки называют бластомерами (от греч. бласте—росток, зачаток). Особенностью митотических делений дробления является то, что с каждым делением клетки становятся все мельче, роста клеток не происходит, митотические циклы в периоде дробления сильно укорочены, особенно в самом начале, следовательно общий размер зародыша к концу дробления на стадии бластулы не превышает размера зиготы. Основные клеточные механизмы. Сначала бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость — бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, образуя стенку бластулы — бластодерму. То есть образуется бластула- многоклеточный односеменной зародыш. Полное (или голобластическое) дробление. Встречается в олиго- и мезолецитальных яйцах. Такой тип дробления встречается у некоторых рыб, всех амфибий, а также у сумчатых и плацентарных млекопитающих. В результате: анимальном полюсе четыре более мелких бластомера — микромеры, на вегетативном — четыре более крупных — макромеры. Затем деление опять идет в меридианных плоскостях, а потом опять в широтных. Частичное (меробластическое) дробление. Встречается в полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих. Оно охватывает только свободную от желтка цитоплазму, которая располагается в виде тонкого диска на анимальном полюсе. При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным. Оно типично для хордовых и иглокожих. Синхронное дробление- образующиеся клетки одинаковы по размеру. Асинхронное дробление-образующиеся клетки различны по размеру. 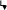 Типы дробления и типы бластул хордовых Типы дробления и типы бластул хордовых Нарушения дробления может произойти при следующих условиях: Изменение свойств жидкости. Например, повышение содержания солей на ранних стадиях может сказаться на форме дробления. Гипертонический раствор – цитоплазма теряет воду, это приводит к повышению её вязкости..итд Повышение температуры. Приводит к повышению скорости дробления. Понижение, наоборот. Наличие в среде кислорода. Отсутствие – дробление не происходит( не у всех). Экстракорпоральное оплодотворение Вспомогательная репродуктивная технология, используемая в случае бесплодия. Во время ЭКО яйцеклетку извлекают из организма женщины и оплодотворяют искусственно в пробирке. Полученный эмбрион содержат в условиях инкубатора, где он развивается в течение 2—5 дней, после чего эмбрион переносят в полость матки для дальнейшего развития. 50.Сущность и способы гаструляции. Гаструляция у различных представителей хордовых. Особенности молекулярно-генетических и биохимических процессов на стадии гаструляции. Нарушения гаструляции. Гаструляция Сущность стадии гаструляции заключается в том, что однослойный зародыш — бластула — превращается в многослойный — двух- или трехслойный, называемый гаструлой. У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок — эктодерму — и внутренний зародышевый листок — энтодерму. Энтодерма формирует первичную кишку с полостью внутри — гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. У позвоночных образуется еще третий зародышевый листок — мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, является эволюционным усложнением фазы гаструляции у позвоночных. У более примитивных хордовых животных, таких, как ланцетник, мезодерма обычно образуется в начале следующей после гаструляции фазы — органогенезе. Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий. Способы гаструляции различны. Выделяют четыре разновидности, приводящих к преобразованию зародыша из однослойного в многослойный. Инвагинация — впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа. Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка. Эпиболия — обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных. Деноминация —расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с частичным типом дробления, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта. Иммиграция — перемещение групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных. Морфология гаструляции. Инвагинация начинается на вегетативном полюсе. Из-за более быстрого деления клетки анимального полюса разрастаются и толкают внутрь бластулы клетки вегетативного полюса. Вследствие инвагинации бластоцель уменьшается, а гастроцель увеличивается. Одновременно с исчезновением бластоцеля эктодерма и энтодерма приходят в тесный контакт. У ланцетника, как и у всех вторичноротых животных область бластопора превращается в хвостовую часть организма в отличие от первичноротых, у которых бластопор соответствует головной части. Ротовое отверстие у вторичноротых образуется на противоположном бластопору конце зародыша. Гаструляция у земноводных имеет много общего с гаструляцией ланцетника, но так как в яйцеклетках у них желтка намного больше и расположен он преимущественно на вегетативном полюсе, крупные бластомеры амфибластулы не способны впячиваться внутрь. Инвагинация проходит иначе. На границе между анимальным и вегетативным полюсами в области серого серпа клетки сначала сильно вытягиваются внутрь, принимая вид «колбовидных», а затем тянут за собой клетки поверхностного слоя бластулы. Возникают серповидная бороздка и спинная губа бластопора. Одновременно более мелкие клетки анимального полюса, делящиеся быстрее, начинают перемещаться в сторону вегетативного полюса. В области спинной губы они подворачиваются и впячиваются, а с боков и со стороны, противоположной серповидной бороздке, обрастают более крупные клетки. Затем образуются боковые и брюшная губы бластопора. Бластопор смыкается в кольцо, внутри которого некоторое время видны крупные светлые клетки вегетативного полюса в виде желточной пробки. Позднее они полностью погружаются внутрь, а бластопор сужается. Конкретные области бластодермы, называемые презумптивными. Известно, что у бесхвостых амфибий материал презумптивной хорды и мезодермы на стадии бластулы лежит не на ее поверхности, а во внутренних слоях стенки амфибластулы. Определенное сходство процессов гаструляции и области презумптивных органов у земноводных и ланцетника, т.е. гомология основных органов, таких, как нервная трубка, хорда, вторичная кишка, указывает на их филогенетическое родство. Гаструляция у зародышей с мepoблacтическим типом дробления и развития имеет свои особенности. К моменту откладки яйца зародыш уже состоит из нескольких слоев: верхний слой называют эпибластом, нижний—первичным гипобластом. Между ними находится узкая щель—бластоцель. Затем образуется вторичный гипобласт. Основные события гаструляции и окончательное образование трех зародышевых листков начинаются после откладки яиц с началом инкубации. Возникает скопление клеток в задней части эпибласта. Образуется первичная полоска, которая вытягивается в направлении к головному концу. В центре первичной полоски образуется первичная бороздка, а по краям—первичные валики. На головном конце первичной полоски возникает утолщение—гензеновский узелок, а в нем —первичная ямка. Когда клетки эпибласта входят в первичную бороздку, их форма изменяется. Они напоминают по форме «колбовидные». Затем эти клетки приобретают звездчатую форму и погружаются под эпибласт, образуя мезодерму. Энтодерма образуется на основе первичного и вторичного гипобласта с добавлением нового поколения энтодермальных клеток, мигрирующих из верхних слоев, бластодермы. |