экзамен магистратура экология. 1. Экология как наука. Место экологии в структуре человеческого знания. В начале xx в сформировалась новая биологическая наука экология. Термин экология
 Скачать 2.4 Mb. Скачать 2.4 Mb.
|

|
13. Отношения типа «хищник – жертва» и «паразит – хозяин» и их краткая характеристика. Отношения типа хищник‑жертва, паразит‑хозяин – это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого – положительные последствия. Подобные популяции развиваются, т. е. эволюционируют, синхронно Размер популяции хищников влияет на размер популяции их жертв и наоборот, динамика популяций описывается математической моделью Лотки - Вольтерра, однако данная модель является высокой степенью абстракции, и не описывает реальные взаимосвязи между хищником и жертвой Колебания системы хищник-жертва (модель Лотки-Вольтера). Еще в 20-х гг. А. Лотка, а несколько позднее независимо от него В.Вольтера предложили математические модели, описывающие сопряженные колебания численности хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтера. Чем может регулироваться численность популяций: Условия (сезонная изменчивость) Ресурсами : конкуренция с особями своего вида Конкуренция с особями других видов Активностью хищников Активностью паразитов и болезней Собственной численностью популяции Модель Лотки-Вольтерры Учитывает изменяющийся пресс хищников при изменении численности их популяций Простая В некоторых отношениях реалистичная Описание взаимодействия популяции жертва и хищник. Модель состоит из двух компонентов: С – численность хищников; N – численность жертв; Предположим, что в отсутствии хищников популяция жертвы будет расти экспоненциально: dN/dt = rN. Но жертвы уничтожаются хищниками со скоростью, которая определяется частотой встреч хищника с жертвой, а частота встреч возрастает по мере увеличения численности хищника (С) и жертвы (N). Точное число встреченных и успешно съеденных жертв будет зависеть от эффективности, с которой хищник находит и ловит жертву, т.е. от а’ – «эффективности поиска» или «частоты нападений». Т.о., частота «успешных» встреч хищника с жертвой и, следовательно, скорость поедания жертв будет равна а’СN и в целом: dN/dt = rN – a’CN (1*). В отсутствии пищи отдельные особи хищника теряют в весе, голодают и гибнут. Предположим, что в рассматриваемой модели численность популяции хищника в отсутствии пищи вследствие голодания будет уменьшаться экспоненциально: dC/dt = - qC, где q – смертность. Гибель компенсируется рождением новых особей со скоростью, которая, как полагают в этой модели, зависит от двух обстоятельств: 1)скорости потребления пищи, a’CN; 2)эффективности (f), с которой эта пища переходит в потомство хищника. Т.о., рождаемость хищника равна fa’CN и в целом: dC/dt = fa’CN – qC (2*). Уравнения 1* и 2* составляют модель Лотки-Вольтера. Свойства этой модели можно исследовать, построить изоклины линии, соответствующие постоянной численности популяции, с помощью таких изоклин определяют поведение взаимодействующих популяций хищник-жертва. В случае с популяцией жертвы: dN/dt = 0, rN = a’CN, или C = r/a’. Т.к. r и a’ = const, изоклиной для жертвы будет линия, для которой величина С является постоянной: При низкой плотности хищника (С), численность жертвы (N) возрастает, наоборот – снижается.  Точно также для хищников (уравнение 2*) при dC/dt = 0, fa’CN = qC, или N = q/fa’, т.е. изоклиной для хищника будет линия, вдоль которой N постоянна: 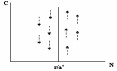 При высокой плотности жертвы, численность популяции хищника растет, а при низкой – снижается. Поместив обе изоклины на 1 рисунок, получим картину взаимодействия популяций:  Их численность претерпевает неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищников на популяцию жертвы и тем самым к уменьшению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и падению их численности, которая вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищников и т.д. Популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новые циклы неограниченных колебаний. На самом деле среда постоянно меняется, и численность популяций будет постоянно смещаться на новый уровень. Чтобы циклы колебаний, которые совершает популяция, были регулярными, они должны быть стабильными: если внешнее воздействие изменяет уровень численности популяций, то они должны стремиться к первоначальному циклу. Такие циклы называются устойчивыми, предельными циклами. Модель Лотки-Вольтера позволяет показать основную тенденцию в отношениях хищник-жертва, которая выражается в возникновении колебаний численности в популяции жертвы, сопровождающихся колебаниями численности в популяции хищника. Основным механизмом таких колебаний является запаздывание во времени, свойственное последовательности состояния от высокой численности жертв к высокой численности хищников, затем к низкой численности жертв и низкой численности хищников, к высокой численности жертв и т.д Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют. Для хищников характерно специальное охотничье поведение. Добыча жертвы требует от них значительных затрат энергии на поиск, погоню, захват, преодоление сопротивления жертв. Если размеры жертв намного меньше размеров питающихся ими животных, численность объектов питания высока и сами они легкодоступны – в этом случае деятельность плотоядного вида превращается в поиск и простой сбор добычи и называется собирательство. Собирательство требует затрат энергии в основном на поиск, а не на захват пищи. Характерно для ряда насекомоядных птиц Во взаимоотношениях хищник‑жертва, паразит‑хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. В процессе совместной эволюции хищники и жертвы приспосабливаются друг к другу (также паразит и хозяин, фитофаг и растение), таким образом достигается динамическое равновесие в системе «хищник-жертва». У хищников появляются и развиваются средства обнаружения и атаки, а у жертв — средства скрытности и защиты. Поэтому наибольший вред жертвам могут нанести новые для них хищники При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы. В свою очередь, трудность обнаружения и поимки жертв способствует у хищников отбору на лучшее развитие органов чувств (зоркость, тонкий слух, чутье и т.д.), на более быструю реакцию на добычу, выносливость при преследовании и т. п. Таким образом, экологические связи хищников и жертв направляют ход эволюции сопряженных видов. Паразитизм - это такая форма пищевой связи между видами, когда организм-потребитель (консумент) использует тело живого хозяина не только как источник пищи, но и как место своего обитания (постоянного или временного). Паразиты намного мельче своего хозяина. Паразитические отношения имеют насекомые-вредители и растения, кровососущие насекомые и животные и т. п. Насекомые-паразиты часто бывают разносчиками эпидемий: вши - тифа, клещи - энцефалита и др. Поскольку хозяин обеспечивает паразиту не только пищу, но и микроклимат, защиту и т. п., то чем лучше приспособленность паразита к особенностям организма хозяина, тем вероятнее его успех в размножении и оставлении потомства. Тесная связь паразита с хозяином имеет следствием отбор двоякого рода. Среди паразитов получают преимущество те, которые способны более полно и длительно использовать хозяина, не приводя его к слишком ранней гибели и обеспечивая тем самым себе наилучшее существование. Иными словами, паразит изнуряет, но не губит хозяина. Катастрофический вред от паразитов выражен преимущественно лишь в тех связях, которые еще не стабилизированы длительным ходом естественного отбора. Поэтому случайно завезенные вредители поражают сельскохозяйственные растения или животных часто намного сильнее, чем местные. Среди многообразных форм паразитических отношений есть и такие, при которых гибель хозяина – обязательное следствие пребывания в нем паразита. Этот тип связей особенно распространен у насекомых, откладывающих свои яйца в яйца или личинки других. Такие насекомые получили названиепаразитоидов. Гибель хозяина обусловлена малым запасом в нем пищи, которой едва хватает на развитие одной или немногих личинок вида-потребителя. Таким образом, при длительном контакте паразитов и хищников с их жертвами, влияние на них весьма умеренно, нейтрально или даже благоприятно, а наиболее повреждающее действие оказывают новые паразиты и хищники. Отсюда вывод: «необходимо избегать создания новых отрицательных взаимодействий, а если они возникли, стараться по возможности сдерживать их» (Ю. Одум, 1975). Основная экологическая роль хищничества, паразитизма и других вариантов пищевых связей в сообществах заключается в том, что, последовательно питаясь друг другом, живые организмы создают условия для круговорота веществ, без которого невозможна жизнь. Вторая не менее важная роль этих отношений – взаимная регуляция численности видов. 14. Гомеостаз и регуляция численности популяций. Понятие популяционной нормы реакции и порогового уровня воздействия. Поддержание определенной численности или равновесное состояние популяции получило название гомеостаза популяции. Гомеостатический механизм – это обратная связь. Например, у пойкилотермных животных изменение температуры тела регулируется специальным центром в мозге, куда постоянно поступает сигнал обратной связи, содержащий данные об отклонении от нормы, а от центра поступает сигнал, возвращающий температуру к норме. Принцип минимального размера популяции состоит в том, что популяция не может состоять из меньшего числа особей, чем необходимо для обеспечения стабильной реализации её структуры и устойчивости к влиянию факторов внешней среды. Правило популяционного максимума -популяция эволюционирует так, что регуляция её плотности осуществляется на значительно более низкой по сравнению с верхней асимптомой емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы Е и пространства. Это правило конкретезируют 2 обобщения: Теория Андреварты – Бирча (1954), или теория лимитов популяционной численности: численность природных популяций ограничена истощением пищевых ресурсов и усл размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе обобщение дополняет первое и носит название теории биоценотической регулции численности популяции К. Фридрикса (1927), регуляции численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида. Экологические системы любого уровня являются гомеостатическими, т.е выполняющими работу по стабилизации среды. Модель популяций как систем, стремящихся стабилизировать свою среду (гомеостатических систем), показывает, что такое стремление может быть реализовано 2 основными способами: Уменьшением смертности с ростом численности и биомассы населения, что соответствует внутрипопуляционной кооперации. Следствием работы механизмов внутрипопуляционной кооперации должно быть увеличение средней продолжительности жизни ( а значит, и «постарение» популяции) с ростом плотности населения. Снижение рождаемости при увеличении плотности – хорошо известное явление внутрипопуляционной конкуренции, которая приводит к стабилизации не только среды, но и самой популяции. Одновременное существование этих основных путей осуществляющихся гомеостатической функции приводит к проявлению принципа Олли – прирост численности мал как при низких, так и при высоких численностях. Концепция авторегуляции численности популяции. Эта концепция базируется на сочетаниях 2 принципиально различных явлений: Модицикации, или случайных колебаний численности. Регуляций, действующих по принципу кибернетической обратной связи и невелирующих эти колебания. В соответствии с этими протипопостовляются- Модифицирующие ( не зависящие от плотности популяций) Регулирующие (зависящие от плотности) -экологические факторы. Модифицирующие факторы могут вызывать весьма значительные отклонения численности популяций в сторону уменьшения или повышения, но реагировать на изменение плотности популяции, т.е действовать по принципу обратной связи, они не могут. К модифицированным факторам, изменяющим, но не регулирующим численность популяции, относятся абиотические факторы. Регулирующие факторы могут не только изменять число особей в популяции, но и регулировать её. Действие этих факторов связано с отрицательной обратной связью с плотностью популяции, т.е интенсивность действия регулирующих факторов, может изменяться под влиянием изменяющейся плотности популяции организмов. К регулирующим факторам относятся биотические факторы – воздействие естественных врагов (хищники, паразиты, возбудители болезней), внутривидовые отношения и пища. 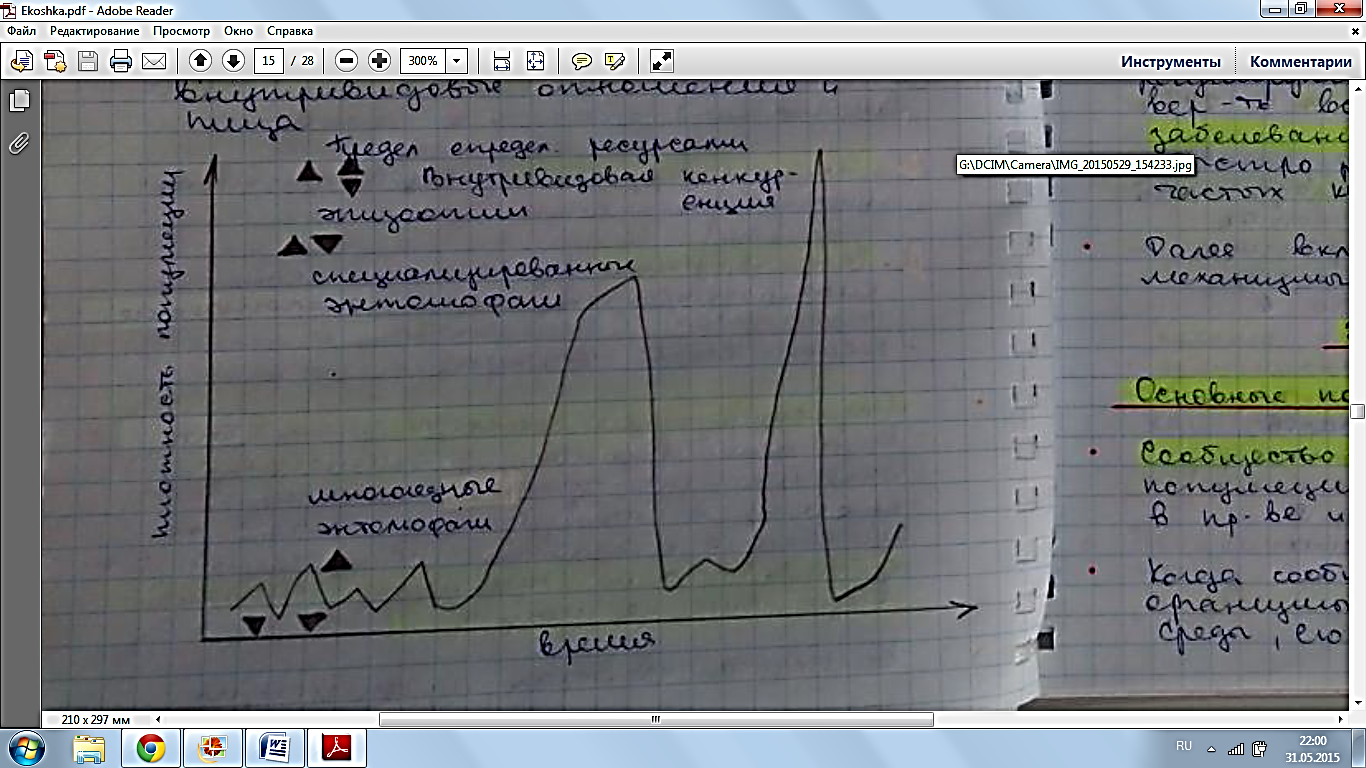 На разных уровнях численности популяции регулирующие факторы принципиально различны. Если темпы роста популяции не очень высоки, то для сдерживания её численности достаточно многоядных хищников. При более быстром увеличении численности хищники теряют регулирующую роль. Однако высокая плотность популяции благоприятна для специализированных паразитов, которым становится легче находить хозяев. При еще более быстром темпе роста популяции паразиты также теряют регулирующую роль, но повышается вероятность вспышки инфекционных заболеваний, тк их возбудители быстро распространяются при частых контактах особей. Далее включаются внутривидовые регулирующие механизмы. Понятие популяционной нормы реакции и порогового уровня воздействия. Любые количественные оценки последствий поступления токсических элементов к животным из природных популяций должны основываться на установленной зависимости «доза-эффект». Двойной характер дозовой зависимости: 1) Градированный вид зависимости. Он в количественной форме отражает изменение того или иного показателя состояния организма как функции внешнего токсического воздействия. Увеличение дозы ведет к подавлению нормального показателя, т.е., нормальное функционирование с увеличением дозы снижается. Градированная зависимость не позволяет сделать заключение о наличии поражения. 2) Альтернативная форма дозовой зависимости подразумевает диагностику состояния «норма-патология». Она основана на концепции пороговости вредного действия загрязнителя. При этом под нормой следует понимать не только совокупность среднестатистических значений основных и важнейших функциональных показателей и параметров, характеризующих состояние организма, но и среднее отклонение от этих значений, связанное как с методическими погрешностями измерения этих параметров, так и в большей степени с индивидуальной изменчивостью рассматриваемых показателей. (Начиная с С активность фермента понижается. С - критическая). Выводы: Для каждого уровня организации биологических систем существуют свои характерные показатели токсичности. Показатели более низких уровней присутствуют в показателях более высокого уровня. Пороговый характер альтернативных эффектов основывается на беспороговости предшествующих показателей. В равной мере беспороговости градированных показателей присутствует альтернативность эффектов предшествующих уровней. Критические значения токсических эффектов сдвигаются в сторону больших значений по мере перехода от низких уровней к более высоким. Популяционный уровень токсических эффектов. Механизмы адаптации. Если равному уровню воздействий подвергать лабораторных организмов и природных популяций, то токсические эффекты, наблюдаемые в эксперименте не проявляются в природе – это явление популяционной адаптации. Явление популяционной адаптации предполагает наличие явления компенсации и явление адаптации. - Компенсация – наличие некоторого уже состоявшегося нарушения биосистемы, которое оказывается компенсированным за счет иных процессов. - Адаптация – приспособление – такая перестройка системы в ответ на длительное токсическое воздействие, которое позволяет этой системе устойчиво сохранять себя и не допускать даже скрытых проявлений токсичности. Например, сущ. перестройки, осуществляемые в пределах нормы: тренированное сердце - увеличивается стенка левого желудочка – адаптация к высоким нагрузкам. При митральном пороке сердца происходит тоже утолщение, но это компенсация к болезни. Существует 2 уровня истинной адаптации: 1) Адаптация на уровне отдельных функциональных систем организма. За счет определенной коррекции биохимических, физиологических или иных реакций мы отмечаем явление повышенной устойчивости. 2) Популяционная адаптация – поддержание популяцией некоторого уровня нормального ее функционирования за счет толерантности (устойчивости) особей, за счет изменения их плодовитости. Эта адаптация базируется на разнокачественности особей, входящих в эту популяцию (гетерогенности популяции). Англичане изучали популяции полевицы на местности, где был установлен медеплавильный завод, происходило аэральное загрязнение, в рез-те загрязнялась почва. Наблюдали в течение 70 лет. Вокруг завода образовался устойчивый покров полевицы (популяционная адаптация). Они проводили корневой тест (определенный индекс толерантности) – отношение средней длины корней растений, культивируемых в питательном растворе с добавками токсичного элемента к средней длине корней растений, культивируемых в питательном растворе без добавок токсического элемента (в чистом растворе). Чем выше корневой индекс, тем более устойчиво раст. N – количество растений, которые обладают определенным индексом толерантности. Инд. – корневой индекс. Чем он больше, тем более толерантно растение. Идея эксперимента: растения, толерантные к Cu были, но их было мало – они не имели преимущества (малоактивно представлены в обычной популяции), но они сильно представлены в условиях загрязнения. Были сделаны работы по птицам-дуплогнездникам. Показатели: величина кладки, размеры яиц, выживаемость птенцов, вылупляемость, весовые данные птенцов перед вылетом из гнезда. В результате непроклюнувшихся яиц в фоновой зоне мало, В буферной зоне – больше, в импактной зоне – очень много (аналогично по другим факторам). В природной популяции все дефекты элиминируют из популяции. Природные популяции исходно гетерогенны, что определяет возможность к адаптации. Норма реакции – спектр возможностей развития в зависимости от наличных условий среды. (определение Яблокова). Комов: возможность развития спектра вариаций признака в определенном диапазоне колебаний внешних условий. Норма реакции женских особей выше, чем у мужских особей. Геоданян: эволюционный смысл увеличения нормы реакции у женских особей: показано на людях (однояйцевые близнецы братья более близки, похожи, чем сестры, так как сестры имеют более широкую норму реакции). Будучи более пластичными, самки способны приспосабливаться лучше, чем менее пластичные самцы. При изменении условий, прежде всего, гибнут самцы, и большем % выживают самки. При смене прир. условий происходит гибель неприспособленных самцов, но всегда есть мизер, который выживает. И этот мизер самцов всегда может покрыть много самок. Ежегодно популяция выбрасывает поколение, много умирает, но есть часть, которая соответствует данным условиям. Есть самки, которые способны далее перенести признаки тех самцов, которые вымерли. (Это свойственно даже для растений). Реагировать на изменения внешней среды организм может только в пределах нормы реакции, заданной программой развития (генотипом). |