экзамен магистратура экология. 1. Экология как наука. Место экологии в структуре человеческого знания. В начале xx в сформировалась новая биологическая наука экология. Термин экология
 Скачать 2.4 Mb. Скачать 2.4 Mb.
|

|
11. Демографическая структура популяции в сообществах. Кривые выживания. Модели роста популяций. Рождаемость и смертность, динамика численности напрямую связаны с возрастной структурой популяции. Популяция состоит из разных по возрасту и полу особей. Для каждого вида, а иногда и для каждой популяции внутри вида характерны свои соотношения возрастных групп. По отношению к популяции обычно выделяют три экологических возраста: предрепродуктивный, репродуктивный и пострепродуктивный. Для многих животных и растений характерен длительный репродуктивный период. У некоторых животных, в особенности у насекомых, пререпродуктивный период чрезвычайно длителен. Репродуктивный период очень короткий, а пострепродуктивный период отсутствует совсем. Возрастные различия в популяции существенно усиливают ее экологическую неоднородность и, следовательно, сопротивляемость среде. Повышается вероятность того, что при сильных отклонениях условий от нормы в популяции сохранится хотя бы часть жизнеспособных особей и она сможет продолжить свое существование. Возрастная структура популяций имеет приспособительный характер. Она формируется на основе биологических свойств вида, но всегда отражает также силу воздействия факторов окружающей среды. Половая структура популяции – это численное соотношение самцов и самок в разных возрастах группах. Половая структура определяется соотношением полов: Первичным (при возникновении зигот); Вторичным (у новорожденных); Третичным (к моменту наступления половой зрелости) Существуют партеногенетические популяции, например, популяция дыбки степной, популяции некоторых видов рыб и рептилий. Другой случай - виды с чередованием поколений. Так, в жизненном цикле живут тлей происходит смена партеногенетических и обоеполых популяций. Возрастная структура популяции характеризует соотношение возрастных групп, отличающихся между собой по отношению к воспроизводству(т.е. какую часть популяции составляют репродуктивные особи). Возрастная структура популяций растений. Периодизация онтогенеза цветковых растений: I Латентный: 1. семена (sm) II Виргинальный: проростки (p) ювенильные (y): молодые растения , по размеру меньше, чем у взрослые особи имматурные (im): переходная форма растений к взрослым вегетативным особям виргинильные (v): хорошо развитые особи растений, но еще не цветущие и не плодоносящие III Генеративный: молодые генеративные (g1) средние генеративные (g2) старые генеративные (g3) IV Сенильный: субсенильные (ss): старые вегетативные растения, характеризующиеся прекращением плодоношения сенильные (s): старые растения, характеризующиеся крайней дряхлостью, уменьшением размеров отмирающие (sc) Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным спектром. Если в возрастном спектре представлены только семена и молодые особи, не достигшие репродуктивной фазы развития, то ценопопуляция называется инвазионной. Она не способна к самоподдержанию. Чаще всего это молодая популяция, только что внедрившаяся в фитоценоз. Если в возрастном спектре представлены все или почти все возрастные группы, то ценопопуляция называется нормальной. Она способна к самоподдержанию генеративным или вегетативным путем. В зависимости от преобладания численности той или иной генеративной группы нормальные ценопопуляции подразделяются на три типа: 1). молодые нормальные - это ценопопуляции, в возрастном спектре которых преобладают молодые генеративные особи; 2). средневозрастные нормальные - это ценопопуляции, в возрастном спектре которых преобладают средневозрастные генеративные особи; 3). старые нормальные - это ценопопуляции, в возрастном спектре которых преобладают старые генеративные особи. Если в возрастном спектре представлены все возрастные группы, то ценопопуляция называется нормальной полночленной, если же некоторые возрастные группы отсутствуют, то она называется нормальной неполночленной. Если ценопопуляция не содержит молодых особей, а представлена только особями сенильными или субсенильными, то такая популяция называется регрессивной. Она не способна к самоподдержанию, это старая популяция. Например, это характерно для многих реликтовых видов секвойи. Возрастную структуру популяций животных изображают в виде возрастных пирамид. В нормальных популяциях должны быть представлены все возрастные группы животных: 1). Ювенильные или молодые особи (juv) 2). Подросшие особи, но ещё не способные к размножению (sad) 3). Взрослые особи с хорошо развитой репродуктивной системой (ad) 4). Сенильные или стареющие особи (sen) В зависимости от продолжительности жизни животных данные возрастные группы могут подразделяться на соответвующие подруппы, например, ad1, ad2 и т.д. Как правило возрастные пирамиды строят для популяции человека в разных странах. Для развитых стран характерна бутылковидная пирамида: высокий уровень медицины. В малоразвитых странах низкая продолжительность жизни, высокая детская смертность, следовательно пирамида второго типа. Популяция могут быть разделены на 3 экологических возраста: пререпродуктивный, репродуктивный и пострепродуктивный. У современного человека 3 этих возраста приблизительно одинаковы. На каждый из них приходится треть жизни. Для многих животных и растений характерен длительный репродуктивный период. У некоторых животных, в особенности у насекомых, пререпродуктивный период чрезвычайно длителен. Репродуктивный период очень короткий, а пострепродуктивный период отсутствует совсем. Возраст созревания Биологическая продукция - баланс прихода вещества и энергии и их затратами на дыхание. Каждая особь может направлять продукцию либо на увеличение своей массы(рост), либо на производство новых членов популяции (рождаемость). "Возраст созревания" - это тот возраст, в котором происходит "переключение" продукции с роста на размножение. В традиционных популяционно-демографических моделях этот возраст жестко фиксирован но сейчас можно считать установленным, что у многих видов животных созревание определяется скорее не возрастом, а размерами. Упрощенно можно принять, что до достижения определенных размеров особи только растут, а после достижения таковых вся продукция направляется на создание новых особей. Кривые выживания. Важнейшим параметром характеристики популяций является выживаемость – число особей ( в пересчете на одну тысячу), достигших определенного возраста. Выживаемость особей описывают кривыми выживания. Для разных видов в одних и тех же условиях эти кривые имеют разную форму, поскольку зависимость уровня смертности от возраста для каждого вида специфична. Различают три типа кривых выживания: К 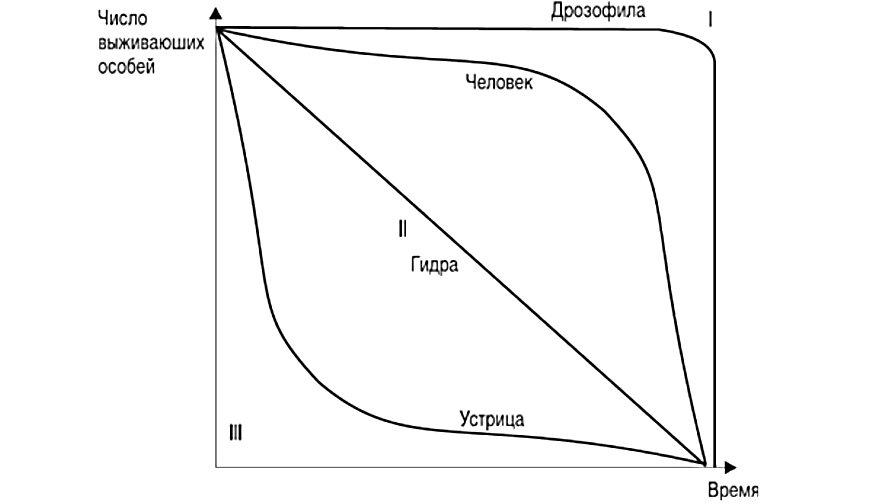 ривая I типа(сильно выпуклая), когда на протяжении всей жизни смертность ничтожно мала, резко возрастая в конце нее, характерна для насекомых, которые обычно гибнут после кладки яиц (ее и называют «кривой дрозофилы»), к ней приближаются кривые выживания человека в развитых странах, а также некоторых крупных млекопитающих. ривая I типа(сильно выпуклая), когда на протяжении всей жизни смертность ничтожно мала, резко возрастая в конце нее, характерна для насекомых, которые обычно гибнут после кладки яиц (ее и называют «кривой дрозофилы»), к ней приближаются кривые выживания человека в развитых странах, а также некоторых крупных млекопитающих.Кривая II типа (диагональная) характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни. Такое распределение смертности не столь уж редкое явление среди организмов. Встречаются они среди рыб, пресмыкающихся, птиц, многолетних травянистых растений. Кривая III типа(сильно вогнутая)- это случаи массовой гибели особей в начальный период жизни. Гидробионты и некоторые другие организмы, незаботящиеся о потомстве, выживают за счет огромного числа личинок, икринок, семян и т. п. Моллюски, прежде чем закрепиться на дне, проходят личиночную стадию в планктоне, где личинки гибнут в огромных количествах, поэтому кривую III называют еще «кривой устрицы». Реальные кривые выживания часто представляют собой некоторую комбинацию указанных выше «основных типов». Например, у крупных млекопитающих, да и у людей, живущих в отсталых странах, кривая I вначале круто падает за счет повышенной смертности сразу после рождения. Модели роста популяций Зависимости скорости роста от плотности называют моделями роста популяций. 1. экспоненциальный рост. Модель предложена Мальтусом в 1798г. Он обратил внимание, что численность популяции растет по экспоненте (в геометрической прогрессии), в то время как производство питания растет со временем линейно (в арифметической прогрессии), из чего сделал справедливый вывод, что рано или поздно экспонента обязательно "обгонит" линейную функцию, и наступит голод. Но ни одна популяция не размножается до бесконечности, должны существовать факторы, препятствующие такому неограниченному размножению. Среди этих факторов может быть нехватка ресурса (продовольствия), вызывающая конкуренцию внутри популяции за ресурс, хищничество, конкуренция c другими видами. Результатом является замедление скорости роста популяции и выход ее численности на стационарный уровень. Н 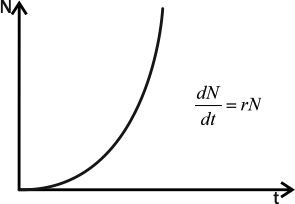 а основании трудов Мальтуса Ч.Дарвин сделал очень важные выводы о том, что поскольку ни одна популяция не размножается до бесконечности, должны существовать факторы, которые препятствуют неограниченному размножению: среди них нехватка ресурсов и конкуренция с другими видами. а основании трудов Мальтуса Ч.Дарвин сделал очень важные выводы о том, что поскольку ни одна популяция не размножается до бесконечности, должны существовать факторы, которые препятствуют неограниченному размножению: среди них нехватка ресурсов и конкуренция с другими видами. N-численность популяции, t- время, где dN/dt - изменение численности популяции (прирост), r- переменная, характеризующая способность популяции увеличивать свою численность(константа) Неограниченный J- образный рост популяции возможен только в среде с неограниченными ресурсами, например, при освоении популяциями новых местообитаний, где ресурсы были накоплены заранее, или в искусственных условиях, где в среду постоянно добавляется пища, и удаляются продукты обмена. Экспоненциальный рост численности популяции – это рост численности ее особей в неизменяющихся условиях. 2.Логистическая модель Ферхюста(S-образная кривая) Ограниченный рост. Описал впервые Ферхюльст в уравнении логистического роста (1848г) dN/dt = r*N*(1- (N/K)) 2 свойства: при малых N численность N возрастает экспоненциально, при больших приближается к определенному пределу K; величина называется емкостью K. 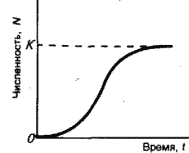 Рост популяций организмов в естественных условиях ограничивается запасами пищи и доступным пространством, пригодным для местообитания - т. е. ёмкостью среды или воздействием лимитирующих (ограничивающих) факторов - сопротивлением среды. Причём эти факторы влияют на численность опосредованно, через изменение системы биотических взаимодействий. Отсюда можно сделать вывод, что существует предел, представляющий собой максимальную численность, к которому будет бесконечно стремиться кривая роста популяции Причины замедления роста популяции могут быть самыми различными: выедание ресурсов, влияние эффекта скученности (у грызунов при этом снижается интенсивность репродуктивного процесса), отравление местообитания прижизненными выделениями, выедание популяции хищниками и т.д. Тем не менее и эта кривая является идеализацией, так как крайне редко проявляется в природе. Очень часто после того, как рост популяции выйдет на плато (достигнет предела К, соответствующего количеству ресурсов), происходит внезапное уменьшение ее численности, а потом популяция вновь быстро растет. Такая циклическая динамика наблюдается, например, в популяциях тундровых леммингов, которые питаются мхами и лишайниками. Они продолжают активную жизнедеятельность под снегом и выедают свою кормовую базу настолько, что прекращают размножаться, а затем начинают умирать от бескормицы. После того, как мхи отрастут, начинается новый подъем численности леммингов. В дифференциальном виде: 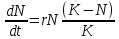 N- числ популяции, r – скорость роста попул. a-const, t – время, К – предельная числ попул. Логистическая модель основывается на очень простых постулатах, которые не выводимы из свойств организмов, однако мы можем себе представить, какими чертами необходимо обладать организмам, чтобы рост их популяции с большей вероятностью описывался логистической кривой. Во-первых, все особи популяции должны быть одинаковыми, т. е. потреблять одно и то же количество пищи (и других ресурсов), при возрастании плотности популяции для них в равной степени должна возрастать вероятность гибели и (или) снижаться вероятность оставить потомство. Во-вторых, реакция этих организмов на возрастание плотности популяции, проявляющаяся в снижении рождаемости и увеличении смертности, должна быть практически мгновенной. Хотя ни один реальный вид организмов такими свойствами не обладает, очевидно, что простейшие или бактерии, т. е. существа мелкие и размножающиеся простым делением, ближе к такому идеалу, чем крупные многоклеточные организмы, характеризующиеся сложным циклом развития и сложной размерно-возрастной структурой популяции. Очевидно также, что, четко осознавая, чем и в какой степени реальные организмы отличаются от идеального объекта логистической модели, исследователь может сделать эту модель более реалистичной, вводя в нее те или иные усложнения, учитывая, например, размерную структуру популяции и эффект запаздывания. Таким образом, на численность популяций влияют самые разнообразные факторы: и погода, и обеспеченность пищей, и хищники, и болезни, и возрастной состав, и соотношение полов и возрастных групп в самой популяции, и многие другие. В этом многообразии, однако, можно четко выделить две группы факторов. Действие одних односторонне. Они влияют на популяцию, но сами не зависят от ее численности и плотности. Таковы в основном все абиотические факторы, например все погодные условия: температура, Дожди, ветры, солнечная радиация, давление и т. п. Они могут обусловить значительную смертность или, наоборот, благоприятствовать размножению. Эти факторы не регулируют плотность популяции, а просто отклоняют ее в ту или иную сторону. Их изучение важно для прогноза численности вида на данной территории. Вторая группа факторов относится к регуляторам численности популяций. Регуляция — это двустороннее взаимодействие. Она возникает по принципу отрицательной обратной связи, когда рост численности популяции вызывает все увеличивающееся противодействие этому росту. Действие регулирующих факторов зависит от плотности популяции. Чем выше становится численность вида, тем сильнее растет противодействие. При падении численности действие регуляторов ослабевает. По этому принципу на популяцию действуют как другие виды, так и рост собственной плотности. Действительно, чем выше численность жертв, тем больше пищи для хищников и паразитов, тем быстрее могут распространяться возбудители опасных заболеваний и тем сильнее обостряется конкуренция внутри собственного вида. Таким образом, факторы, регулирующие численность популяции, т. е. возвращающие ее к норме из состояния переуплотнения, — это в основном межвидовые и внутривидовые отношения, т. е. биотические связи. Именно они удерживают плотность популяции в определенных границах, не допуская виды до критического состояния — подрыва собственных ресурсов. 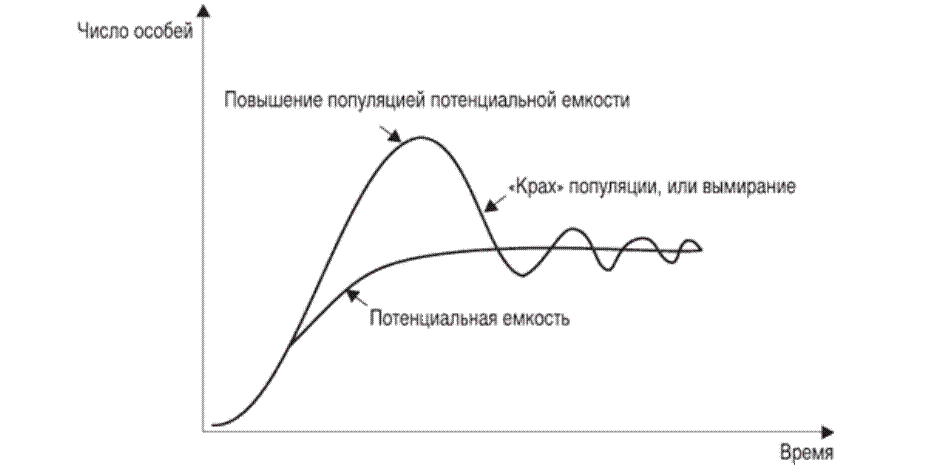 Кроме того, емкость среды может меняться как в зависимости от сезона, так и в зависимости от года, в результате в популяции наблюдаются нерегулярные колебания численности, так как каждый раз проскок численности происходит относительного другого значения К. Такие нерегулярные колебания численности называются флуктуациями .(пример китобойный промысел) Кроме того, емкость среды может меняться как в зависимости от сезона, так и в зависимости от года, в результате в популяции наблюдаются нерегулярные колебания численности, так как каждый раз проскок численности происходит относительного другого значения К. Такие нерегулярные колебания численности называются флуктуациями .(пример китобойный промысел)В природе различают сезонные и годичные флуктуации. В свою очередь годичные флуктуации можно разделить на две группы: 1) флуктуации, контролируемые годичными различиями внешних факторов; 2) флуктуации, связанные с самой динамикой популяции. Из всех причин, вызывающих колебания численности, некоторые исследователи главную роль отводят климатическим условиям, считая биотические факторы второстепенными. По мнению других, только те факторы, которые зависят от плотности, вызывают колебания численности. Многие ученые признают роль тех и других. В экосистемах с низким уровнем видового разнообразия численность популяций подвержена сильному воздействию физических стрессоров и зависит от погоды, химического состава среды и степени ее загрязнения. В экосистемах с высоким уровнем видового разнообразия колебания численности популяций в основном контролируются биотическими факторами. 12. Взаимоотношения популяций в сообществах. Классификация, характеристика основных типов взаимоотношений. Теоретически взаимодействие популяций двух видов можно выразить в виде следующих комбинаций символов: 00, --, ++, +0, -0 , +-. Выделяют 9 типов наиболее важных взаимодействий между видами (по Ю. Одуму, 1986): Примечание: (0) - существенное взаимодействие между популяциями отсутствует; (+) - благоприятное действие на рост, выживание или другие характеристики популяции; ( - ) - ингибирующее действие на рост или другие характеристики популяции. * нейтрализм(00)- ассоциация двух видов популяций не сказывается ни на одном из них; * Взаимное конкурентное подавление (--) - обе популяции взаимно подавляют друг друга; * Конкуренция из-за ресурсов (--) - каждая популяция неблагоприятно воздействует на другую при недостатке пищевых ресурсов; * Аменсализм (-0) - одна популяция подавляет другую, но сама при этом не испытывает отрицательного влияния; * Паразитизм (+ - ) -популяция паразита наносит вред популяции хозяина; * Хищничество (+ -) - одна популяция неблагоприятно воздействует на другую в результате прямого нападения, но зависит от другой; * Комменсализм (+0) - одна популяция извлекает пользу от объединения с другой, а другой популяции это объединение безразлично; * Протокооперация (+ +) -обе популяции получают пользу от объединения; * Мутуализм (+ +) - связь благоприятна для роста и выживания отдельных популяций, причём в естественных условиях ни одна из них не может существовать без другой. Девять описанных видов взаимодействий можно свести к двум более обобщенным типам - отрицательным (антибиотическим) и положительным (симбиотическим). Далее на примерах подробнее рассмотрим типы взаимоотношений между организмами. В случае, когда одна популяция не испытывает влияния другой, т.е. между ними нет взаимодействия, то такая ситуация называется нейтрализмом. Например, белки и лоси в одном лесу не контактируют между собой. К антибиотическим отношениям можно отнести следующие формы отношений: * Конкуренцию; * Паразитизм; * Хищничество; * Аменсализм. Если популяции являются антагонистами в борьбе за пищу, место обитания и другие необходимые для жизни факторы, то их отношения называют конкуренцией. Конкуренция – это соревнование организмов одного трофического уровня (м\у растениями, м\у фитофагами и т д) за потребление ресурсов, имеющихся в ограниченном количестве. Одно живое существо лишает части ресурса другое, которое вследствие этого медленнее растет, оставляет меньшее число потомков и имеет больше шансов погибнуть. Лишать друг друга потенциального ресурса могут особи как одного, так и разных видов. В первом случае взаимное влияние можно считать равноценным и симметричным, так как особи одного вида имеют более сходные потребности в ресурсах, чем особи разных видов. К. делится на межвидовую и внутривидовую. Конкуренция: за пищу - Гиена и стервятники; Антилопа гну и зебра; песец и поморник. за территорию - медведи труться о деревья, оставляя метки, пение соловья - сигнал, что территория занята, В природе возникают ситуации, когда один вид наносит ущерб другому, но в то же время не может существовать без него. Такой тип взаимоотношений называют либо паразитизмом (когда представители какого-либо вида обитают внутри или на поверхности другого - например, паразитические черви, обитающие внутри млекопитающих и человека), либо хищничеством (когда представители одного вида ловят и поедают представителей другого - например, отношения между волками и грызунами и др.). При аменсализме страдает один вид, а другой развивается нормально: например, плесневый гриб Penicillium выделяет пенициллин - вещество, подавляющее рост различных бактерий, но бактерии не оказывают влияния на плесневый гриб. При антогонизме - аменсализме, паразитизме и хищничестве - ошибочно полагать, что отношения между хозяином и паразитом или хищником и жертвой всегда вредны для хозяина или жертвы. Со временем под влиянием естественного отбора губительное действие паразита или хищника ослабевает, поскольку паразит может сохраниться лишь в том случае, если перейдет на какой-либо новый вид, пригодный в качестве хозяина. Если же паразит не найдет нового хозяина, то он погибает сам. К симбиотическим можно отнести следующие формы отношений: * Собственно симбиоз (протокооперация); * Мутуализм; * Комменсализм. Симбиоз - сожительство представляет такую форму взаимоотношений, при которой оба партнера или один из них извлекает пользу от другого. Симбиоз - понятие широкое и включает много разных вариантов отношений между видами. Если присутствие одной популяции благоприятно для другой, но не является необходимым условием существования, то такие отношения носят характер протокооперации (на панцирях многих ракообразных обитают различные кишечнополостные, которые получают пищу, когда его хозяин ловит и поедает других животных, однако они могут существовать и раздельно). Протокооперацию можно в определенном смысле отнести к симбиозу. Существуют и другие формы симбиоза: мутуализм, комменсализм. В случае, когда оба вида извлекают выгоду из совместного существования и не могут жить самостоятельно, то такая ассоциация называется мутуализмом. Примером мутуализма являются термиты, в кишечнике которых обитают жгутиковые (простейшие), имеющие ферменты для разложения древесины, которой питаются термиты. Формирование мутуализма проходит через несколько стадий: сначала ассоциация носит характер комменсализма (т.е. такой тип взаимоотношений, когда один из двух совместно обитающих видов - комменсал - извлекает пользу из совместного существования, не причиняя, однако, вреда другому виду), а затем через фазу протокооперации отношения в ассоциации переходят в мутуализм. Комменсализм широко представлен в океане, где практически в каждой норе, вырытой червем, и в каждой раковине обитают гости, использующие убежище хозяина и не приносящие ему ни пользы, ни вреда. Рыбы-лоцманы, следующие за акулами, черепахами, дельфинами, кормятся остатками пищи этих животных, а также их экскрементами и паразитами. Такие отношения между видами называют нахлебничеством (один из вариантов комменсализма). Другая форма комменсализма получила название квартиранства: в полости голотурии "морского огурца" находят убежище разнообразные мелкие виды животных. Растения - эпифиты (от греч. "эпи" - на, сверх, "фитом" - растение) поселяются на деревьях. Например, на деревьях поселяются водоросли, лишайники, мхи, орхидеи - они питаются за счет фотосинтеза и отмирающих тканей хозяина, но не их соками. |