экзамен магистратура экология. 1. Экология как наука. Место экологии в структуре человеческого знания. В начале xx в сформировалась новая биологическая наука экология. Термин экология
 Скачать 2.4 Mb. Скачать 2.4 Mb.
|

|
15. Экологические стратегии выживания, r- и К-типы. Система типов стратегий Раменского – Грайма. В экологии и эволюционной биологии для описания способов выживания популяций и их роли в сообществе в условиях совместного использования необходимых ресурсов получили широкое распространение два типа репродуктивных стратегий, связанные с энергетическими затратами на поддержание продолжительности жизни и на процессы размножения: r– и k–стратегии. Эти стратегии разработаны Макартуром и Уильсоном, как результаты двух типов отбора r-отбор и k-отбор. r-стратегия-высокая смертность при высокой рождаемости K-стратегия-низкая смертность при низкой рождаемости r-отбор - эволюция в направлении увеличения затрат на размножение организма, итогом которой являются r-стратегии К-отбор - эволюция в направлении увеличения затрат на поддержание жизни взрослого организма, ее итогом являются К-стратеги. r-стратегия – ею обладают быстро размножающиеся виды (r-виды); для нее характерен отбор на повышение скорости роста популяции в периоды низкой плотности. Она характерна для популяций в среде с резкими и непредсказуемыми изменениями условий или в эфемерных, т.е. существующих короткое время (пересыхающие лужи, заливные луга, временные водотоки) Основные признаки r-видов: высокая плодовитость, короткое время регенерации, высокая численность, обычно малые размеры особей (у растений мелкие семена), малая продолжительность жизни, большие траты энергии на размножение, кратковременность местообитаний, низкая конкурентоспособность. R-виды быстро и в больших количествах заселяют не занятые территории, но, как правило, скоро – в течение жизни одного-двух поколений сменяются К-видами. К r-видам относятся бактерии, все однолетние растения (сорняки) и насекомые-вредители (тли, листоеды, стволовые вредители, стадная фаза саранчи). Из многолетников – пионерные виды: Иван-чай, многие злаки, полыни, эфемерные растения, из древесных видов – ивы, березы белая и каменная, осина, чозения, из хвойных – лиственница; они появляются первыми на нарушенных землях: гарях, горных полигонах, строительных карьерах, по обочинам дорог. K-стратегия – этой стратегией обладают виды с низкой скоростью размножения и высокой выживаемостью (К-виды); она определяет отбор на повышение выживаемости при высокой плотности популяции, приближающейся к предельной. Основные признаки К-видов: низкая плодовитость, значительная продолжительность жизни, крупные размеры особей и семян, мощные корневые системы, высокая конкурентоспособность, устойчивость на занимаемой территории, высокая специализация образа жизни. Скорость размножения К-видов с приближением к предельной плотности популяции падает и быстро увеличивается при низкой плотности; родители заботятся о потомках. К-виды часто становятся доминантами биогеоценозов. К К-видам относятся все хищники, человек, реликтовые насекомые (крупные тропические бабочки, в т.ч. дальневосточные, реликтовый усач, жук-олень, жужелицы и др.), одиночная фаза саранчи, почти все деревья и кустарники. Наиболее яркие представители растений – все хвойные, дуб монгольский, орех маньчжурский, лещины, клены, разнотравье, осоки. Р 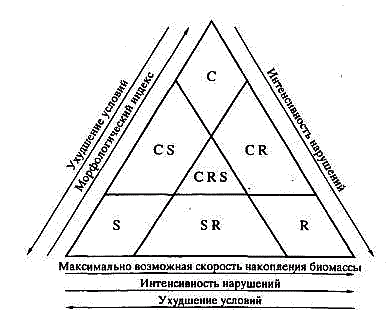 азные популяции по-разному используют одну и ту же среду обитания, поэтому в ней одновременно могут существовать виды обоих типов стратегией. азные популяции по-разному используют одну и ту же среду обитания, поэтому в ней одновременно могут существовать виды обоих типов стратегией. Система типов стратегий Раменского-Грайма. Рис.треугольник Грайма. Отношение к факторам: 1. Обеспеченность ресурсами. 2. Противодействие стрессу. С-конкурентные- малые нарушения, хорошие условия. Береза. S-стресс-толлеранты- Высокий стресс, но малые нарушения. Бук. R-рудералы- однолетние травы. Высокий стресс и высокий уровень нарушений. Первичные типы стратегий. C-стратеги (competitor - конкурент)=виоленты= львы. Это мощные организмы с затрачиванием энергии на поддержание жизни взрослого организма. В основном деревья, иногда кустарники, травы, наподобие тростника. Требуют хороших условий отсутствия нарушений. Виоленты в равной степени неустойчивы как к ухудшению условий (просыхание почвы, засоление и т.д.), так и к нарушениям (рубка леса). Под воздействием этих факторов виоленты, как правило, погибают, так как лишены специальных приспособлений для поддержания устойчивости в таких условиях. Чистый виолент – редкость, чаще встречаются вторичные типы, переходные от виолента к другим типам стратегий. Мб распростертая крона, широкие корни и тд. R - стратеги (ruderis сорный) = эксплерент = шакал. Используют ресурсы в то время, когда они не востребованы, например, львами. Эти организмы замещают виолетнов при сильных нарушениях местообитаний. Представлены в основном травянистым ярусом. . Эти растения замещают виоленты при сильных нарушениях местообитаний (истинные эксплеренты) или используют ресурсы в стабильных местообитаниях, но в период, когда они оказываются невостребованными доминантами (так называемые ложные эксплеренты). Большинство эксплерентов – однолетники или реже малолетники с высоким энергетическими расходами на размножение (репродуктивным усилием). Марь, лебеда, пижма. S-стратеги (stress - tolerant) = патиент = верблюд. Засуха, засоление и тд. Это разнообразные организмы, способные за счет специальных адаптаций переживать сильный стресс. Растения патиенты обитают при дефиците ресурсов или при наличии условий, которые ограничивают их потребление (засуха, засоление, дефицит света или ресурсом минерального питания, холодный климат и т.д.). Они могут обитать под пологом виолентов. Вторичные типы стратегий. Многие виды способны совмещать разные типы стратегий. Сосна относится к категории CS, так как она хорошо растет на бедных песчаных почвах; CR- ольха серая, ольховник, крапива- так как она доминирует на нарушенных местообитаниях; SR-некоторые однолетние травы (щучка). Третичные. CRS – степные, луговые травы. Более подробные классификации выделяют и другие, промежуточные типы. В частности, можно различать еще группу пионерных видов, которые быстро занимают вновь возникающие территории, на которых еще не было никакой растительности. Пионерные виды частично обладают свойствами эксплерентов – низкой конкурентной способностью, но, как и патиенты, обладают высокой выносливостью к физическим условиям среды. В 70-е годы прошлого века, спустя 40 лет после Л. Г. Раменского, выделение тех же трех фитоценотипов повторил незнакомый с его классификацией ботаник Д. Грайм, обозначив их другими терминами: конкуренты, толеранты и рудералы. 16. Устойчивость экосистем. Основные положения. Одним из важнейших показателей динамики экосистем является устойчивость. Определение устойчивости экосистемы - это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия. Также следует различать два вида устойчивости: резидентная устойчивость (стабильность) - способность оставаться в устойчивом (равновесном) состоянии под нагрузкой, и упругая устойчивость (собственно устойчивость) - способность быстро восстанавливаться при снятии нагрузки. Длительность существования каждой экосистемы поддерживается прежде всего за счет общего круговорота веществ, осуществляемого продуцентами, консументами и редуцентами, и постоянного притока солнечной энергии. Именно эти два глобальных явления обеспечивают ей высокую способность противостоять воздействию постоянно меняющихся условий внешней среды. Устойчивость экосистемы обеспечивается также биологическим разнообразием и сложностью трофических связей организмов, входящих в ее состав. В богатых видами экосистемах у консументов есть возможность избирать разные виды пищевых объектов и в первую очередь — наиболее массовые. Если потребляемый пищевой объект становится редким, то консумент переключается на питание другим видом, а первый, освобожденный от пресса выедания, постепенно будет восстанавливать свою численность. Благодаря такому переключению поддерживается динамическое равновесие между пищевыми ресурсами и их потребителями и обеспечивается возможность их длительного сосуществования. Важным фактором стабилизации экосистемы является генетическое разнообразие особей популяций. Изменение условий внешней среды может вызвать гибель большинства особей популяции, адаптированных к прежним условиям существования. Поэтому чем более генетически разнородной является та или иная популяция экосистемы, тем больший шанс у нее иметь организмы с аллелями, ответственными за появление признаков и свойств, позволяющих выжить и размножаться в новых условиях и восстановить прежнюю численность популяции. Время, необходимое для восстановления популяции, будет зависеть от скорости размножения особей, так как изменение признаков происходит только путем отбора в каждом поколении. Стабильность экосистемы зависит также от степени колебаний условий внешней среды. В тропиках и субтропиках стабильны и оптимальны для многих видов температурные условия, влажность, освещенность. Поэтому тропические экосистемы с высоким биологическим разнообразием входящих в них организмов отличаются высокой устойчивостью. И, напротив, тундровые экосистемы менее устойчивы. Им свойственны резкие колебания численности популяций разных видов. Следует различать: Упругая экосистема способна воспринимать значительные воздействия, не изменяя существенно своей структуры и свойств. Однако, при определенной интенсивности воздействий она обычно разрушается или переходит в качественно новое состояние. Пластичная экосистема - более чувствительна к внешним воздействиям, но под их влиянием она "прогибается". После прекращения воздействия относительно быстро возвращается в исходное или близкое к нему состояние. Иногда выделяют третий аспект устойчивости — устойчивость экосистемы по отношению к изменениям характеристик среды и изменению своих внутренних характеристик. В случае, если экосистема устойчиво функционирует в широком диапазоне параметров окружающей среды и/или в экосистеме присутствует большое число взаимозаменяемых видов (то есть, когда различные виды, сходные по экологическим функциям в экосистеме, могут замещать друг друга), такое сообщество называют динамически прочным (устойчивым). В обратном случае, когда экосистема может существовать в весьма ограниченном наборе параметров окружающей среды, и/или большинство видов незаменимы в своих функциях, такое сообщество называется динамически хрупким (неустойчивым) Закон Эшби: Стабильность и устойчивость экосистемы тем выше, чем больше видовое разнообразие. Для экосистемы с низкой устойчивостью характерны вспышки численности отдельных видов. Способность экосистемы к саморегуляции и поддержанию динамического равновесия называется гомеостазам. Гомеостаз экосистемы выражается в способности сохранять постоянство видового состава и численности особей, поддерживать относительную стабильность и целостность генетической структуры в меняющихся условиях внешней среды. Нарушение природных цепей питания под воздействием антропогенного фактора, непродуманное вмешательство человека в экосистемы могут привести к неконтролируемому росту или снижению численности особей определенных популяций и к нарушению природных экосистем. Устойчивость тем значительнее, чем разнообразнее экосистемы. Тундровые и пустынные экосистемы рассматриваются как малоустойчивые (нестабильные), а тропические леса, максимально богатые по видовому составу, - как самые устойчивые (стабильные). Для экосистем с низкой устойчивостью характерны вспышки численности отдельных видов, т.к. в маловидовых экосистемах слабо проявляются силы, уравновешивающие численность различных видов (конкуренция, хищничество, паразитизм). Для тундровых экосистем типичны периодические резкие увеличения численности мелких грызунов - леммингов. К неустойчивым и низкостабильным относят агросистемы, создаваемые человеком и представленные обычно одним преобладающим видом растений, интересующим человека. Устойчивость, стабильность и другие параметры экосистем зависят часто не столько от структуры самих сообществ (например, их разнообразия), сколько от биолого-экологических свойств видов-эдификаторов и доминантов, слагающих эти сообщества. Основные положения экосистемы: устойчивость круговоротов, биоразнообразие, равновесие между тремя уровнями(продуценты, консументы, редуцент). Закон Линдермана, с первого уровня может переходить 10% энергии, если больше, то устойчивость нарушается, происходит деградация, переход на низший уровень(луг- опустынивание). 17. Продуктивность и биологическая продукция. Особенности проявления продуктивности по поверхности Земли. Продуктивность экосистемы – это скорость, с которой продуценты усваивают лучистую энергию в процессе фотосинтеза и хемосинтеза, образуя органическое вещество. Биологическая продукция – скорость накопления биомассы в экосистеме, отражающая способность организмов производить органическое вещество в процессе своей жизнедеятельности. Биологическая продукция измеряется количеством органического вещества, создаваемого за единицу времени на единицу площади (m/ Га/ год, кг/ кВ.м/год, г/кв.м/день и т.д. ) Различают разные уровни продуцирования, на которых создается первичная и вторичная продукция. Скорость, с которой продуценты экосистемы фиксируют солнечную энергию в химических связях синтезируемого органического вещества, определяет продуктивность сообществ. Органическую массу, создаваемую растениями за единицу времени, называют первичной продукцией сообщества. Продукцию выражают количественно в сырой или сухой массе растений либо в энергетических единицах – эквивалентном числе джоулей. Первичная продукция подразделяется - валовую и чистую продукцию. Валовая первичная продукция – количество вещества, создаваемого растениями за единицу времени при данной скорости фотосинтеза. Часть этой продукции идет на поддержание жизнедеятельности самих растений (траты на дыхание). Эта часть может быть достаточно большой. В тропических лесах и зрелых лесах умеренного пояса она составляет от 40 до 70 % валовой продукции. Планктонные водоросли используют на метаболизм около 40 % фиксируемой энергии. Оставшаяся часть созданной органической массы характеризует чистую первичную продукцию, которая представляет собой величину прироста растений. Чистая первичная продукция – это энергетический резерв для консументов и редуцентов. Перерабатываясь в цепях питания, она идет на пополнение массы гетеротрофных организмов. Прирост за единицу времени массы консументов – это вторичная продукция сообщества. Вторичная продукция не делится уже на валовую и чистую, так как консументы и редуценты, т. е. все гетеротрофы, увеличивают свою массу за счет первичной продукции, т. е. используют ранее созданную продукцию. Вторичную продукцию вычисляют отдельно для каждого трофического уровня, так как прирост массы на каждом из них происходит за счет энергии, поступающей с предыдущего. Все живые компоненты экосистемы - продуценты, консументы и редуценты - составляют общую биомассу («живой вес») сообщества в целом или его отдельных частей, тех или иных групп организмов. Биомассу обычно выражают через сырой и сухой вес, но можно выражать и в энергетических единицах - в калориях, джоулях и т. п., что позволяет выявить связь между величиной поступающей энергии и, например, средней биомассой. На образование биомассы расходуется не вся энергия, но та энергия, которая используется, создает первичную продукцию и может расходоваться в разных экосистемах по-разному. Если скорость ее изъятия консументами отстает от скорости прироста растений, то это ведет к постепенному приросту биомассы продуцентов и возникает избыток мертвого органического вещества. Последнее приводит к заторфовыванию болот, зарастанию мелких водоемов, созданию большого запаса подстилки в таежных лесах и т. п. В стабильных сообществах практически вся продукция тратится в трофических сетях, и биомасса остается постоянной. По первичной биологической продукции (в сухом веществе, Р Уиттекер, 1980) различают экосистемы на 4 класса: 1. Очень высокая (>2 кг/м2 – в год) характерна для влажных тропических лесов, коралловых рифов, геотермальных «оазисов» рифтовых зон океана, плавней – зарослей тростника в дельтах великих рек. 2. Высокая (1- 2 кг/м2 в год) – широколиственные леса, прибрежные заросли тростника на озере, посевы кукурузы и многолетних трав. 3. Умеренная (0,25 – 1 кг/м2 в год) преобладающая часть сельхозугодий, бореальные леса, сенокосные луга и степи, заросшие озера, «морские луга» из водорослей. 4. Низкая (<0,25 кг/м2 в год) – настоящие и арктические пустыни, тундры, полупустыни Прикаспия, вытоптанные скотом степные пастбища, каменистые степи. Большинство морских экосистем зоны пела Средняя биологическая продукция экосистем Земли не превышает 0,3 кг/м2 в год, т.к. на планете преобладают низкопродуктивные экосистемы. Потребленная пища = рост + дыхание + фекалии + экскреты Энергия, теряющаяся при дыхании, не передается другим организмам. Энергия, заключенная в экскрементах и отходах метаболизма, передается детритофагам и редуцентам, она не теряется для экосистемы. Детритные цепи начинаются с мертвых организмов и отмерших частей растений. Доля чистой первичной продукции, переходящей в детрит и в пищевые цепи редуцентов, неодинакова в разных системах. В лесной экосистеме большая часть первичной продукции поступает в детритные цепи, а не в пастбищные. Поэтому лесная подстилка оказывается местом значительной активности консументов. Но в морских экосистемах и на интенсивно используемых пастбищах больше половины первичной продукции может поступать в пастбищную пищевую цепь. Если экосистема стабильна и в ней не происходит увеличения общей биомассы, то биомасса в конце года будет такая же, как и в начале. Вся энергия, заключенная в первичной продукции, пройдет через различные трофические уровни, а чистая продукция будет равна нулю. Но обычно система изменяется. Например, лес к концу вегетационного периода накапливает часть поступившей энергии в форме прироста биомассы. При расчете продуктивности учитывают сезонные изменения продуктивности за год. Продуцирование – непрерывный процесс, поэтому при расчетах общей продукции организмов необходимо учитывать приросты и выживших, и погибших в течение данного промежутка времени. В сообществе, сохраняющем устойчивое состояние, фактическая продукция данного трофического уровня должна покрывать пищевые потребности организмов следующего уровня. Продукция, как и энергия, убывает резко при переходе от низших трофических уровней к высшим. Ежегодная продукция зависит от видового состава, географической широты и высотной поясности: * Влажные тропики – 300-500 ц/га; * Широколиственные леса – 80-150 ц/га; * Горные луга – 120 ц/га; * Тайга – 85-40 ц/га; * Степи – 13-50 ц/га; * Пустыни – 5-15 ц/га (бореальные, полярные и т.п.); * Верховые болота – 22 ц/га; * Тундра – 5 ц/га; * Каменистые пустыни <1ц/га |