1. Физиология наука о жизнедеятельности организма, его взаимодействия с окружающей средой и динамикой жизненных процессов. Значимость современной физиологии и её связь с другими науками
 Скачать 0.8 Mb. Скачать 0.8 Mb.
|

|
12. Законы раздражения возбудимых тканей. Эти законы отражают определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани. К законам раздражения относятся: закон силы, закон «все или ничего», закон раздражения Дюбуа-Реймона (аккомодации), закон силы-времени (силы-длительности), закон полярного действия постоянного тока, закон физиологического электротона. Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционирует скелетная мышца. Амплитуда ее сокращений постепенно увеличивается с увеличением силы раздражителя вплоть до достижения максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), на пороговые раздражители возникает максимальная ответная реакция («все»). По закону «все или ничего» сокращаются сердечная мышца и одиночное мышечное волокно. Закон «все или ничего» не абсолютен. Во- первых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в ткани происходят изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, реагирует по закону «все или ничего», но амплитуда ее сокращения будет больше по сравнению с таковой при сокращении нерастянутой сердечной мышцы. Закон раздражения Дюбуа-Реймона (аккомодации): стимулирующее действие постоянного тока зависит не только от абсолютной величины силы тока, но и от скорости нарастания тока во времени. При действии медленно нарастающего тока возбуждение не возникает, так как происходит приспособление возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при дейст-вии медленно нарастающего раздражителя в мембране происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения ПД не возникает. Закон силы-времени: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать на возбудимые ткани, чтобы вызвать возбуждение. Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании — под анодом. Прохождение постоянного электрического тока че- рез нервное или мышечное волокно вызывает изменение мембранного потенциала. Так, в области приложения катода положительный потенциал на наружной стороне мембраны уменьшается, возникает деполяризация, которая быстро достигает критического уровня и вызывает возбуждение. В области же приложения анода положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает. Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом и в соседних с ним участках понижается вследствие деполяризации мембраны (возбудимость повышается). В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости). Повышение возбудимости под катодам называется катэлектротоном, а снижение возбудимости под ано- дом — анэлектротоном. 13. Парабиоз. Свойства парабиотического возбуждения. Функциональный парабиоз. Оптимум и пессимум. Парабиоз. Возбудимость может быть снижена или полностью устранена действием ряда факторов. Одни из них блокируют мембранные ионные каналы (нейротоксины, наркотики, двух- и трехвалентные катионы). Другие вызывают устойчивую длительную деполяризацию мембраны, во время которой она абсолютно невозбудима. Такую стойкую деполяризацию Н.Е. Введенский рассматривал как особую форму «застойного» нераспространяющегося возбуждения. Свойства парабиотического возбуждения. Парабиотическое возбуждение характеризуется рядом особенностей: • ограничено областью нанесения раздражения, т.е. стационарно и не распространяется по возбудимой ткани; • непрерывно; • возбудимость парабиотического участка противоположна возбудимости неповрежденного соседнего участка; • область парабиоза характеризуется электроотрицательностью мембраны, изменяющейся по мере углубления парабиоза. Функциональный парабиоз. Оптимум и пессимум. Сильные и частые раздражения вызывают столь серьезные изменения в биологических структурах, что это не может не отразиться на их функциях. В основе этих изменений лежит явление функционального парабиоза. На нервно-мышечном препарате установлено, что сильные и частые раздражения нерва электрическим током ведут к ослаблению или полному прекращению мышечных сокращений. Это состояние называется пессимумом физиологической реакции. Уменьшение силы и частоты раздражения приводит к увеличению амплитуды сокращения — оптимуму физиологической реакции. Механизм оптимума и пессимум а. В происхождении пессимума важную роль играет длительность абсолютного, и в меньшей степени относительного, рефрактерных периодов, а в явлении оптимума — продолжительность фазы супернормальной возбудимости. • Если каждый последующий раздражающий импульс «попадает» в период супернормальной возбудимости предшествующего цикла возбуждения, то конечная физиологическая реакция возрастает до оптимальной величины (оптимум реакции). • Если каждый последующий импульс раздражающего тока совпадает с периодом абсолютной рефрактерности предыдущего цикла возбуждения, новый цикл возбуждения не возникает. Следовательно, длительность периода абсолютной рефрактерности является критической количественной мерой лабильности, своеобразной «мертвой зоной», разделяющей следующие потенциалы действия и придающей импульсации дискретный характер. • Если каждый последующий раздражающий импульс «попадает» в период относительной рефрактерности предыдущего цикла, возбуждение может иметь местный характер и не вызовет физиологической реакции, или вызовет — но только в том случае, если будет значительно превышать пороговое значение. Продолжительность рефрактерной фазы и фазы супернормальной возбудимости меняется в процессе стимуляции. При увеличении суммарного интервала абсолютной и относительной рефрактерной фаз пессимальная реакция выражена резче. 14. Физиологические свойства и функции скелетной мышцы. Структурно-функциональные основы скелетной мышцы. Микроструктура актинового и миозинового филаментов. Свойства: 1)возбудимость – способность отвечать на раздражения возникновением ПД; 2)проводимость – способность к проведению ПД вдоль мышечного волокна; 3)сократимость – способность изменять длину и напряжение при возбуждении; 4)растяжимость; 5)эластичность Функции: Передвижение тела в пространстве; Перемещение частей тела друг относительно друга; Поддержание позы; Передвижение крови и лимфы; Выработка тепла; Участие в акте вдоха и выдоха; Двигательная активность; Депонирование воды и солей; Защита внутренних органов Физиологические свойства: Входят в состав опорно-двигательного аппарата; Имеют быструю кратковременную деполяризацию и короткий период абсолютной рефрактерности; Не обладают способностью к дифференцировке и делению; Иннервируются соматической нервной системой; Сокращаются под влиянием импульсов, передаваемых по двигательным нервам от мотонейронов спинного мозга; Способны к быстрым фазическим сокращениям; Не имеют пластического тонуса; Осуществляют произвольные мышечные движения, сопровождаемые значительными энергетическими затратами; Обладают слабо выраженной чувствительностью к химическим веществам; В незначительной степени управляемы лекарственными средствами Мышца состоит из мышечное волокон, которые состоят из миофибрилл, включающих саркомеры(структурно-функциональная ед-ца миофибриллы), состоящих из актиновых и миозиновых филаментов( Анизотропный диск, инизотропный диск и т-системы) Актиновый филамент: цепь тропомиозина с насаженым на нее белком актином и тропонином, включающим 3 субъединицы. Субъединица Т-связывает актин, субъединица C- связывает кальций и субъединица I-ингибирует связь актин-миозин, подавляет гидролиз АТФ. Миозиновый филамент: два толстого конца миозинового филамента состоят из множества молекул миозина. Хвосты тяжелой цепи миозина(2шт) скручены на подобие рулета и связаны за счет электростатических сил. Головка располагается в виде поперечных мостиков благодаря чему она может дотягиваться до нити актина, связываться с ней и сокращаться. На месте перехода головки в хвост располагаются легкие цепи (4шт), оказывающие влияние на способность связывания актина с миозином. 15. Функциональная характеристика гладких мышц. Функции и основные физиологические свойства. Механизм сокращения гладкой мышцы. Основные физиологические свойства: Формируют оболочки внутренних органов и сосудов; Имеют медленную деполяризацию и длительный период абсолютной рефрактерности; Обладают способностью к дифференцировке, делению, регенерации при повреждении; Иннервируются ВНС , а также имеют автономный базовый аппарат иннервации; Сокращаются под влиянием импульсов, передаваемых по вегетативным нервам, а также возникающих в самих мышках; Способны к длительным тоническим сокращениям; Имеют пластический тонус; Осуществляют непроизвольные мышечные сокращения, сопровождаемые незначительными энергетическими потерями; Обладают высокой чувствительностью к химическим, фармакологическим, эндогенным и экзогенным биологически активным веществам; В значительной степени управляемы лекарственными веществами. Функции: Обеспечивают функцию полых органов, стенки которых они образуют; Осуществляют изгнание содержимого из мочевого пузыря, кишки и т.д.; Обеспечивают сфинктерную функцию (создают условия для хранения содержимого полого органа в этом органе); Играют роль в системе кровообращения и лимфообращения; Могут влиять на функцию связочного аппарата и при своем сокращении менять состояние данной связочной структуры. Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре. При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам. Сокращение и расслабление гладких мышц . Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам. Сокращение гладкомышечной клетки: Сокращение гладкомышечной клетки возникает в ответ на развитие в ней ПД, который увеличивает вход ионов кальция через мембрану клетки и их освобождение из внутриклеточных депо. Ионы кальция вместе с кальций- связывающим белком кальмодулином активируют фермент (киназу легких цепей миозина), который переносит фосфатную группу с АИФ на миозин, вызывая взаимодействие актина и миозина, т.е. сокращение мышцы. Расслабление происходит, если концентрация ионов кальция ниже 8-10 моль/л. Но в связи со слабым развитием саркоплазматического ретикулума расслабление происходит медленнее, чем у поперечнополосатых мышц. 16. Молекулярный механизм сокращения скелетной мышцы (теория «скользящих нитей»). В основе сокращения мышц (саркомеров) лежит взаимное перемещение двух систем нитей, образованных актином и миозином. Сами актиновые и миозиновые нити своей длины не изменяют. АТФ гидролизуется в активном центре, расположенном в головках миозина. Гидролиз сопровождается изменением ориентации головок миозина и перемещением нитей актина к середине саркомера. Функция сокращения обеспечивается специальными Са – связывающими белками (тропонин и тропомиозин). В основе сокращения лежит скольжение тонких миозиновых нитей относительно толстых актиновых. При сокращении не меняется длина нитей, но увеличивается область их перекрывания. В процессе сокращения происходит уменьшение ширины J- диска и увеличение А – диска за счёт уменьшения ширины Н -полоски. При расслаблении происходит обратное увеличение Н – полоски, увеличение J-диска. Показано, что молекула миозина образует так называемые миозиновые мостики. Каждый состоит из головки и шейки. Молекула актина обладает сродством к головке миозинового мостика. Суть сокращения состоит в том, что головка миозинового мостика связывается с актином, затем происходит сокращение шейки. В дальнейшем связь актина и миозина разрывается и головка перескакивает на последующий мостик, обеспечивая процесс скольжения нитей. Замечено, что головки одной половины толстого миозинового филамента ориентированы в сторону одной Z -мембраны, а второй половины – в противоположную сторону, к другой Z-мембране. На головке миозинового мостика имеется 4 последовательно рас- положенных центра связывания М1-М4. Сродство к актину возрастает от М1 к М4. В результате поворот головки происходит только в одну сторону. При этом растягивается шейка и нарастает сила сокращения (рис. 27). Энергообеспечение сокращения связано с энергозатратами со- крашения и разрыва связей актина и миозина. Актин обладают АТФ- азной активностью. АТФ-азная активность миозина слабая. Она возрастает на несколько порядков при образовании актино-миозинового комплекса. Для разрыва связи актина с головкой миозинового мостика необходимы энергия АТФ и магний. После разрыва связи головка перескакивает дальше по молекуле актина. 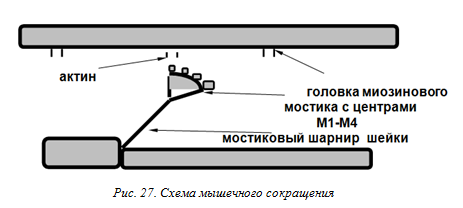 |