1 Генетика, предмет и задачи. Понятие о наследственности и изменчивости. Генетика
 Скачать 0.61 Mb. Скачать 0.61 Mb.
|

|
F: [В одном эксперименте самца, имеющего определенный доминантный признак, скрещивают с самкой, имеющий рецессивный признак. Во втором, соответственно, скрещивают самку с доминантным признаком и самца с рецессивным признаком. Используется для определения роли пола в наследовании признака, а также позволяет определить, от какого из родителей передаются потомству цитоплазматические наследственные факторы. Для проведения реципрокного скрещивания родительские формы должны быть чистыми линиями. В настоящее время, в связи с развитием молекулярной генетики, практическое значение реципрокного скрещивания снизилось.] 11) Дигибридное скрещивание. III закон Г. Менделя. [Скрещивание, при котором родительские формы отличаются по двум парам альтернативных признаков, называется дигибридным. Если родители отличаются по многим парам альтернативных признаков, скрещивание называется полигибридным. Дигибридное скрещивание, по сути, представляет собой два независимо идущих моногибридных скрещивания. Закон независимого наследования (третий закон Менделя) — при скрещивании двух особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). В основе независимого наследования лежит: 1) случайное расхождение негомологичных хромосом в анафазе I мейоза, которое приводит к формированию гамет с различными комбинациями генов; 2) случайное слияние гамет при оплодотворении, что обусловливает образование разных типов зигот. Когда скрещивались гомозиготные растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам, и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Гены, контролирующие развитие разных пар признаков, называются неаллельными. Они располагаются в разных парах хромосом или в разных участках гомологичных хромосом. В данном случае гены, обусловливающие окраску, неаллельны по отношению к генам, определяющим поверхность семян. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.] 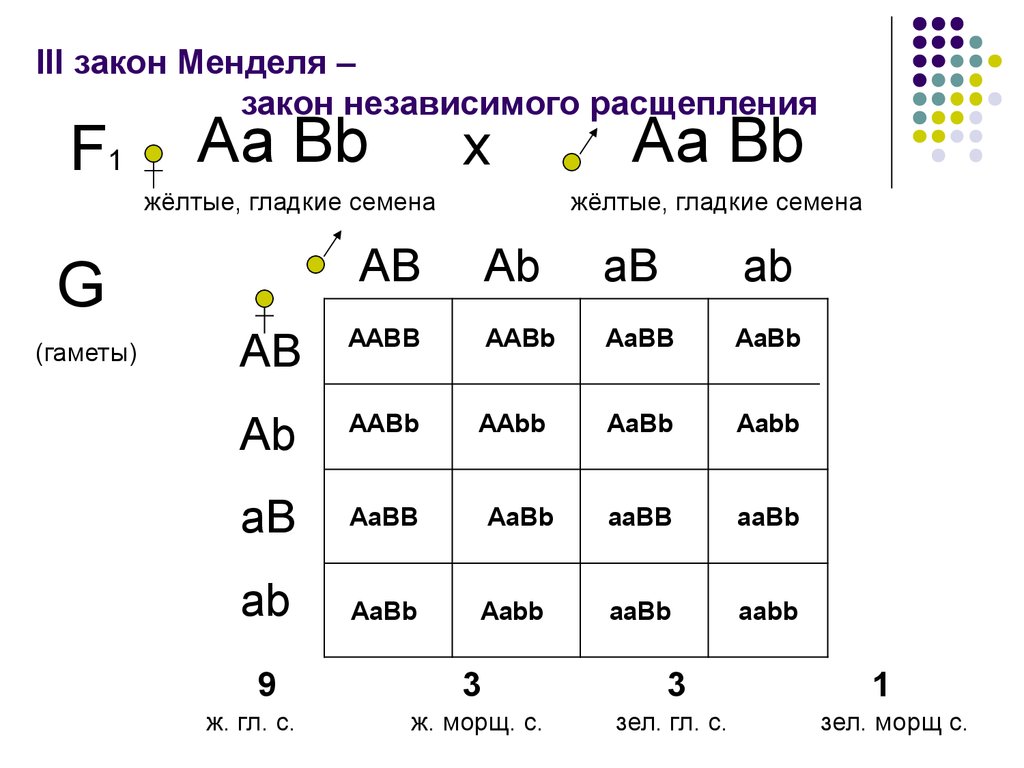 12. Цитологические основы дигибридного скрещивания. Дигибридное скрещивание – скрещивание особей, отличающихся между собой по двум парам альтернативных признаков. Результаты дигибридного скрещивания зависят от того, лежат ли гены, в одной хромосоме или в разных. При дигибридном скрещивании Мендель изучал наследование признаков, за которые отвечают гены, лежащие в разных хромосомах. Грегор Мендель изучал независимое наследование признаков у гороха. Он скрещивал сорта гороха, которые отличались друг от друга двумя парами признаков. В момент переоткрытия закона Менделя цитология накопила достаточно знаний о развитии полов клеток. Была проведена закономерность расщепления в хромосомах при мейозе. В процессе мейоза у гибридных организмов в анафазе осуществлялось расхождение к полюсам гомологичных хромосом каждой пары, при этом негомологичные хромосомы у полюсов комбинировались случайно в возможных сочетаниях. Хромосома несущая аллель А с равной вероятностью может отойти к одному полюсу, имеющая аллель В, так и аллель в. 13. Дигибридное анализирующее скрещивание. Анализирующее скрещивание – скрещивание гибридной особи с особью гомозиготной по рецессивным аллелям. При полном доминировании, среди особей с доминантными признаками невозможно отличить гомозиготы от гетерозиготы, а в этом часто бывает необходимость, например, чтобы опредилить, чистопородна или гибридна данная особь. С этой целью проводят анализирующее скрещивание, при котором исследуемая особь с доминантными признаками (генотип которой неизвестен) скрещивается с рецессивной гомозиготой – генотип которой известен. Если особь, обладающая доминантными признаками, гетерозиготна по двум парам признакам, то в потомстве наблюдается расщепление в соотношении 1:1:1:1. Р ♀ АаВв х ♂ аавв G АB; Аb; аВ; аb; аb
14. Тригибридное скрещивание. Тригибридным называют такое скрещивание, когда родительские формы различаются по трем парам альтернативных признаков. Расщепление будет равно 27: 9: 9: 9: 3: 3: 3: 1. Примером может служить скрещивание двух сортов гороха: один с желтыми гладкими семенами и пурпурной окраской цветков, другой – с зелеными и морщинистыми семенами и белой окраской цветков. У тригибридных растений проявятся доминантные признаки – желтая окраска и гладкая форма семян, а также пурпурная окраска цветка (закон единообразия гибридов F1). РР ♀ААВВСС х ♂♂ ааввсс гаметы: АВС авс F1 АаВвСс Тригибридное растение в результате независимого расщепления генов продуцирует 8 типов гамет (АВС, авс, АВс, авС, Авс, аВС, аВс, АвС) женских и мужских. 15. Взаимодействие неаллельных генов: комплементарность. Комплементарное взаимодействие – действие генов, заключающееся в том, что фенотип у особи развивается только в том случае, еси в ее генотипе присутствие одновременно неаллельного гена находится в гомо- или гетерозиготном состоянии. 2 типа комплем-ти: 1) Такое взаимодействие при котором ни один из доминантных генов не имеет фенотипического выражения. D – улитка d – глухота E – слухов нерв е – глухота D_E_ - норм D_e_ - глухота 2) Взаимодействие генов, при котором один из доминантных генов проявляется фенотипически, а другой неаллельный доминантный ген модифицирует его действие. D – пигмент d – белый E – агути (ген-модификатор) е – белый D_E_ - агути D_eе – пигментированный ddE_ - белые ddee – белые 9:3:4 по фенотипу 16. Взаимодействие неаллельных генов: эпистаз. Эпистаз – один из типов взаимодействия неаллельных генов, сущность которого сводится к подавлению действия доминантного гена одной пары аллели генами другой пары. Ген, подавляющий фенотипические проявления другого, называется эпистатичным. 2 типа: Доминантный (А>B) Рецессивный (a>B) P: AABB (сер) x aabb (рыж) F1: AaBb (сер) F2: 12 сер, 3 вороных, 1 рыжая АВ – сер а, в – рыжие А – серая В – вороная А>В 9 А_В_ - серая 3 А_вв – серая 3 ааВ_ - вороная 1 аавв – рыжая 17. Взаимодействие неаллельных генов: полимерия. Полимерия – наследование при котором один признак находится под контролем нескольких пар генов с однонаправленным действием. 2 типа: Кумулятивная. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Некумулятивная. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Пример полимерии — наследование цвета кожи у людей, который зависит от четырёх генов с кумулятивным эффектом. 18. Структурно-функциональная организация хромосом. Строение хромосом. Хромосомы - это интенсивно окрашенное тельце, состоящее из молекулы ДНК, связанной с белками-гистонами. Хромосомы формируются из хроматина в начале деления клеток (в профазе митоза), но лучше их изучать в метафазе митоза. Хромосомы состоят из 2 сестринских хроматид (удвоенных молекул ДНК), соединенных друг с другом в области первичной перетяжки - центромеры. Центромера делит хромосому на 2 плеча. Классификация хромосом по расположения центромеры: 1) метацентрические (плечи одинакового размера) 2) субметацентрические (одни короткие, другие длинные) 3) акроцентрические (хромосомы, центромера которых находится на конце или второе плечо настолько мало, что малоразличимо на цитологических препаратах) 4) телоцентрические (центромера находится в конце) Центромера = первичная перетяжка. Она определяет движение хромосомы. Имеет сложное строение. На хромосоме только одна центромера. На хромосоме имеются вторичные перетяжки (ядрышковые организаторы), которые не являются местом прикрепления нитей веретена деления. Но они определяют угол изгиба хромосомы при движении. Стр-ра хромосом, видимых в световой микроскоп: 1) темные участки (гетерохроматин) 2) светлые участки (эухроматин) Спутник – участки, соед с хромосомой, поср-вом тонкой нити хроматина и функциональной ответственности не несут (наход у акро- типа) Дифференционная окраска хромосом: 1) Q и H окраска. Флуоресцениные красители. Они позволяют выявлять участки хромосом, обогащенные парами А и Т 2) G окраска. Выявляют участки объед генами, содерж повышенное кол-во Аи Т 3) R окраска. Участки, обогащены генами которые содержат повышенное число пар Г и Ц 4) Т окраска. Вариант, предназначенный для выявления преимущественно концевых теломерных участков, обогащенных Г и Ц. 19. Упаковка ДНК в хромосомах. 1) Нуклеосомный – на этом уровне двойная спираль ДНК наматывается на белковый комплекс, содержащий 8 молекул гистонов – белков с повышенным содержанием положительно заряженных аминокислотных остатков лизина и аргинина. Нуклеосомный уровень даёт укорочение молекулы ДНК в 7 раз. Ведет к сверхскручиванию ДНК 2) Нуклеомерный. Этот уровень упаковки, как и первый, не зависит от первичной структуры ДНК. 6 нуклеосом объединяются в глобулу. 3) Хромомерный = Петлевой уровень. Обеспечивается негистоновыми белками. Они узнают определённые последовательности ДНК и связываются с ними и друг с другом, образуя петли по 20-80 тыс. п.н. 4) Хромонемный. Происходит сближение хромомеров с последующим образованием толстых нитей. Хар-р упаковки выглядит как спиральная уклодки петлевых стр-р. 20. Кариотип. Идеограмма. Постоянные характеристики хромосомного набора, их число и морфологические особенности, наблюдаемые при помощи диффер окрасти, используют для описания кариотипа. Кариотип – признаки, которые в комплексе позволяют идентифицировать данный хромосомный набор, то есть число хромосом, их форма, особенности расположения центромеры, чередование гетеро- и эухроматиновых районов. Кариотип введен в 1924 году Левитским. В кариотипе присутствуют аутосомы (пары хромосом, свойственные представителям обоих полов) и одна пара половых хромосом. Число хромосом у некоторых объектов генетики: Человек – 46 Свинья – 38 Курица – 78 Мышь – 40 Дрозофила – 8 Сазан - 104 Папоротник – 1200 Кариотип может быть изображен в виде идеограммы. На ней изображается по одной из пары гомологичных хромосом. Хромосомы расположены по мере убывания их величины. На идеограмме изображаются не только морфол. признаки хромосом, но и особенности их первичной структуры, спирализации, р-ны гетерохроматина и др. 21. Клеточный цикл. Клеточный цикл – промежуток жизни клетки от ее образования до ее деления да 2 дочерние. Промежуток времени от одного деления клетки до другого называется интерфазой. У разных организмов и в разных тканях продолжительность клеточного цикла различна. Клеточный цикл соматических клеток высших растений и животных можно разделить на 2 стадии: митоз и интерфаза. Существуют 2 способа деления клеток: - митоз (см вопр 22) - мейоз (см вопр 23) Интерфазу делят на 3 периода: G1 пресинтетический, или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов; S синтетический, во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть). G2 постсинтетический, во время которой идет подготовка к митозу. 22. Митоз, фазы и значение. Митоз – непрямое деление соматических клеток, в результате которого из одной диплоидной клетки получается 2 диплоидные точные копии материнских. 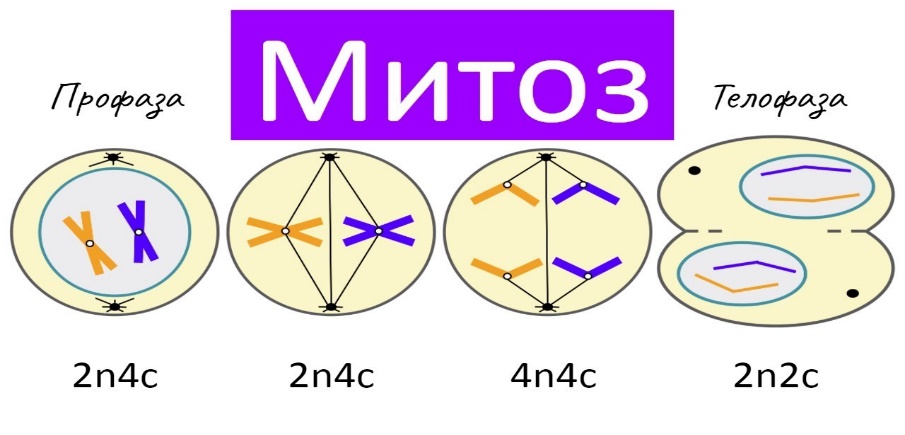 Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу. В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид и формируется веретено деления. Постепенно растворяются ядрышки и ядерная оболочка. Содержание генетического материала в клетке остается неизменным (2n4c). В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки. Содержание генетического материала не изменяется (2n4c). В анафазе каждая хромосома «расщепляется» на две хроматиды, которые с этого момента называются дочерними хромосомами. Нити веретена сокращаются и тянут хроматиды (дочерние хромосомы) к противоположным полюсам клетки. Содержание генетического материала в клетке у каждого полюса представлено диплоидным набором хромосом, но каждая хромосома содержит одну хроматиду (4n4c). В телофазе расположившиеся у полюсов хромосомы деспирализуются и становятся плохо видимыми. Вокруг хромосом у каждого полюса формируется ядерная оболочка, в ядрах образуются ядрышки. Разрушается веретено деления. Дочерние клетки имеют диплоидный набор хромосом, каждая из которых состоит из одной хроматиды (2n2c). Биологическое значение митоза. Оно состоит в том, что митоз обеспечивает наследственную передачу признаков и свойств в ряду поколений клеток при развитии многоклеточного организма. Благодаря точному и равномерному распределению хромосом при митозе все клетки единого организма генетически одинаковы. Митотическое деление клеток лежит в основе всех форм бесполого размножения как у одноклеточных, так и у многоклеточных организмов. Митоз обусловливает важнейшие явления жизнедеятельности: рост, развитие и восстановление тканей и органов и бесполое размножение организмов. 23. Мейоз, фазы и значение. Мейоз – способ деления, в результате которого из диплоидной клетки образуется гаплоидная. Состоит из 2-х стадий Мейоз 1 и Мейоз 2. Каждый делится на 4 фазы (профазу, метафазу, анафазу и телофазу). В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр). В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр). В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое. Содержание генетического материала становится 1n2хр у каждого полюса. В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр). Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза. В профазе мейоза II происходят те же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp). Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Биологическое значение мейоза: 1) является основным этапом гаметогенеза; 2) обеспечивает передачу генетической информации от организма к организму при половом размножении; 3) дочерние клетки генетически не идентичны материнской и между собой. |