1. Медицинская микробиология. Предмет, задачи, методы, связь с другими науками. Значение медицинской микробиологии в практической деятельности врача
 Скачать 0.96 Mb. Скачать 0.96 Mb.
|

|
Иммуноглобулин класса G. Изотип G состав ляет основную массу Ig сыворотки крови. На его долю приходится 70—80 % всех сывороточ ных Ig, при этом 50 % содержится в тканевой жидкости. Среднее содержание IgG в сыворот ке крови здорового взрослого человека 12 г/л. Период полураспада IgG — 21 день. IgG — мономер, имеет 2 антигенсвязывающих центра (может одновременно свя зать 2 молекулы антигена, следовательно, его валентность равна 2), молекулярную массу около 160 кДа и константу седиментации 7S. Различают подтипы Gl, G2, G3 и G4. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо опре деляется в сыворотке крови на пике первич ного и при вторичном иммунном ответе. Обладает высокой аффинностью. IgGl и IgG3 связывают комплемент, причем G3 ак тивнее, чем Gl. IgG4, подобно IgE, обладает цитофильностью (тропностью, или сродс твом, к тучным клеткам и базофилам) и участ вует в развитии аллергической реакции I типа. В иммунодиагностических реакциях IgG может проявлять себя как не полное антитело. Легко проходит через плацентарный барь ер и обеспечивает гуморальный иммунитет новорожденного в первые 3—4 месяца жизни. Способен также выделяться в секрет слизис тых, в том числе в молоко путем диффузии. IgG обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществля ет запуск комплемент-опосредованного цито лиза и антителозависимой клеточно-опосредованной цитотоксичности. Иммуноглобулин класса М. Наиболее круп ная молекула из всех Ig. Это пентамер, кото рый имеет 10 антигенсвязывающих центров, т. е. его валентность равна 10. Молекулярная масса его около 900 кДа, константа седи ментации 19S. Различают подтипы Ml и М2. Тяжелые цепи молекулы IgM в отличие от других изотипов построены из 5 доменов. Период полураспада IgM — 5 дней. На его долю приходится около 5—10 % всех сывороточных Ig. Среднее содержание IgM в сыворотке крови здорового взрослого человека составляет около 1 г/л. Этот уровень у человека достигается уже к 2—4-летнему возрасту. IgM филогенетически — наиболее древний иммуноглобулин. Синтезируется предшест венниками и зрелыми В-лимфоцитами. Образуется в начале первичного иммунного ответа, также первым начинает синтезиро ваться в организме новорожденного — опре деляется уже на 20-й неделе внутриутробного развития. Обладает высокой авидностью, наиболее эффективный активатор комплемента по клас сическому пути. Участвует в формировании сывороточного и секреторного гуморального иммунитета. Являясь полимерной молекулой, содержащей J-цепь, может образовывать сек реторную форму и выделяться в секрет сли зистых, в том числе в молоко. Большая часть нормальных антител и изоагглютининов относится к IgM. Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыво ротке крови новорожденного указывает на бывшую внутриутробную инфекцию или де фект плаценты. IgM обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществля ет запуск комплемент-опосредованного цито лиза и антителозависимой клеточно-опосредованной цитотоксичности. Иммуноглобулин класса А. Существует в сы вороточной и секреторной формах. Около 60 % всех IgA содержится в секретах слизистых. Сывороточный IgA:На его долю прихо дится около 10—15% всех сывороточных Ig. В сыворотке крови здорового взрослого чело века содержится около 2,5 г/л IgA, максимум достигается к 10-летнему возрасту. Период полураспада IgA — 6 дней. IgA — мономер, имеет 2 антигенсвязывающих центра (т. е. 2-валентный), молекуляр ную массу около 170 кДа и константу седи ментации 7S. Различают подтипы А1 и А2. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо опре деляется в сыворотке крови на пике первич ного и при вторичном иммунном ответе. Обладает высокой аффинностью. Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентар ный барьер. IgA обеспечивает нейтрализацию, опсони-зацию и маркирование антигена, осуществля ет запуск антителозависимой клеточно-опос-редованной цитотоксичности. Секреторный IgA:В отличие от сывороточ ного, секреторный sIgA существует в полимерной форме в виде ди- или тримера (4- или 6-валентный) и содержит J- и S-пeптиды. Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше. Синтезируется зрелыми В-лимфоцитами и их по томками — плазматическими клетками со ответствующей специализации только в пре делах слизистых и выделяется в их секреты. Объем продукции может достигать 5 г в сутки. Пул slgA считается самым многочисленным в организме — его количество превышает суммарное содержание IgM и IgG. В сыворотке крови не обнаруживается. Секреторная форма IgA — основной фак тор специфического гуморального местного иммунитета слизистых оболочек желудочно-кишечного тракта, мочеполовой системы и респираторного тракта. Благодаря S-цепи он устойчив к действию протеаз. slgA не активи рует комплемент, но эффективно связывается с антигенами и нейтрализует их. Он препятс твует адгезии микробов на эпителиальных клетках и генерализации инфекции в преде лах слизистых. Иммуноглобулин класса Е. Называют так же реагином. Содержание в сыворотке крови крайне невысоко — примерно 0,00025 г/л. Обнаружение требует применения специаль ных высокочувствительных методов диагнос тики. Молекулярная масса — около 190 кДа, константа седиментации — примерно 8S, мо номер. На его долю приходится около 0,002 % всех циркулирующих Ig. Этот уровень дости гается к 10—15 годам жизни. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками преиму щественно в лимфоидной ткани бронхолегочного дерева и ЖКТ. Не связывает комплемент. Не проходит че рез плацентарный барьер. Обладает выражен ной цитофильностью — тропностью к тучным клеткам и базофилам. Участвует в развитии гиперчувствительности немедленного типа — реакция I типа. Иммуноглобулин класса D. Сведений об Ig данного изотипа не так много. Практически полностью содержится в сыворотке крови в концентрации около 0,03 г/л (около 0,2 % от общего числа циркулирующих Ig). IgD имеет молекулярную массу 160 кДа и константу се диментации 7S, мономер. Не связывает комплемент. Не проходит че рез плацентарный барьер. Является рецепто ром предшественников В-лимфоцитов. Природа иммуноглобулинов. В ответ на введение антигена иммунная систе ма вырабатывает антитела — белки, способные специфически со единяться с антигеном, вызвавшим их образование, и таким образом участвовать в иммунологических реакциях. Относятся ан титела к γ-глобулинам, т. е. наименее подвижной в электричес ком поле фракции белков сыворотки крови. В организме γ-глобулины вырабатываются особыми клетками — плазмоцитами. γ-глобулины, несущие функции антител, получили название иммуноглобули нов и обозначаются символом Ig. Следовательно, антитела — это иммуноглобулины, вырабатываемые в ответ на введение анти гена и способные специфически взаимодействовать с этим же антигеном. Функции. Первичная функция состоит во взаимодсйствии их активных центров с комплементарными им де терминантами антигенов. Вторичная функция состоит в их способности: • связывать антиген с целью его нейтрализации и элиминации из организма, т. е. принимать участие в формировании защи ты от антигена; • участвовать в распознавании «чужого» антигена; • обеспечивать кооперацию иммунокомпетентных клеток (мак рофагов, Т- и В-лимфоцитов); • участвовать в различных формах иммунного ответа (фагоци тоз, киллерная функция, ГНТ, ГЗТ, иммунологическая то лерантность, иммунологическая память). Структура антител. Белки иммуноглобулинов по химическому составу относятся к гликопротеидам, так как состоят из проте ина и Сахаров; построены из 18 аминокислот. Имеют видовые отличия, связанные главным образом с набором аминокислот. Их молекулы имеют цилиндрическую форму, они видны в электронном микроскопе. До 80 % иммуноглобулинов имеют константу седиментации 7S; устойчивы к слабым кисло там, щелочам, нагреванию до 60 °С. Выделить иммуноглобули ны из сыворотки крови можно физическими и химическими ме тодами (электрофорез, изоэлектрическое осаждение спиртом и кислотами, высаливание, аффинная хроматография и др.). Эти методы используют в производстве при приготовлении иммуно биологических препаратов. Иммуноглобулины по структуре, антигенным и иммунобио логическим свойствам разделяются на пять классов: IgM, IgG, IgA, IgE, IgD. Иммуноглобулины М, G, А имеют под классы. Например, IgG имеет четыре подкласса (IgG,, IgG2, IgG3, IgG4). Все классы и подклассы различаются по аминокис лотной последовательности. Молекулы иммуноглобулинов всех пяти классов состоят из полипептидных цепей: двух одинаковых тяжелых цепей Н и двух одинаковых легких цепей — L, соединенных между собой дисульфидными мостиками. Соответственно каждому классу иммуноглобулинов, т.е. М, G, A, E, D, разли чают пять типов тяжелых цепей: μ (мю), γ (гамма), α (альфа), ε (эпсилон) и Δ (дельта), различающихся по антигенности. Легкие цепи всех пяти классов являются общими и бывают двух типов: κ (каппа) и λ (ламбда); L-цепи иммуноглобулинов различных классов могут вступать в соединение (рекомбинироваться) как с гомологичны ми, так и с гетерологичными Н-цепями. Однако в одной и той же молекуле могут быть только идентичные L-цепи (κ или λ). Как в Н-, так и в L-цепях имеется вариабельная — V область, в которой последовательность амино кислот непостоянна, и константная — С область с постоянным набором аминокислот. В легких и тяжелых цепях различают NH2- и СООН-концевые группы. При обработке γ -глобулина меркаптоэтанолом разрушаются дисульфидные связи и молекула иммуноглобулина распадается на отдельные цепи полипептидов. При воздействии протеолитическим ферментом папаином иммуноглобулин расщепляется на три фрагмента: два не кристаллизующихся, содержащих детерминантные группы к антигену и названных Fab-фрагментами I и II и один кристаллизующий Fc-фрагмент. FabI- и FabII-фрагменты сходны по свойствам и аминокислотному составу и отличаются от Fc-фрагмента; Fab-и Fc-фрагменты являются компактными образованиями, соеди ненными между собой гибкими участками Н-цепи, благодаря чему молекулы иммуноглобулина имеют гибкую структуру. Как Н-цепи, так и L-цепи имеют отдельные, линейно свя занные компактные участки, названные доменами; в Н-цепи их по 4, а в L-цепи — по 2. Активные центры, или детерминанты, которые формиру ются в V-областях, занимают примерно 2 % поверхности мо лекулы иммуноглобулина. В каждой молекуле имеются две де терминанты, относящиеся к гипервариабельным участкам Н-и L-цепей, т. е. каждая молекула иммуноглобулина может свя зать две молекулы антигена. Поэтому антитела являются двух валентными. Типовой структурой молекулы иммуноглобулина является IgG. Остальные классы иммуноглобулинов отличаются от IgG дополнительными элементами организации их молеку лы. В ответ на введение любого антигена могут вырабатываться антитела всех пяти классов. Обычно вначале вырабатывается IgM, затем IgG, остальные — несколько позже. первичный и вторичный от вет. Способность к образованию ан тител появляется во внутриутробном периоде у 20-недельного эмбриона; после рождения начинается собственная продукция иммуноглобулинов, которая увеличивается до наступления зре лого возраста и несколько снижается к старости. Динамика об разования антител имеет различный характер в зависимости от силы антигенного воздействия (дозы антигена), частоты воздействия антигена, состояния организма и его иммунной системы. При первичном и повторном введении антигена динамика антителообразования также различна и протекает в несколько ста дий. Выделяют латентную, логарифмическую, стацио нарную фазу и фазу снижения. 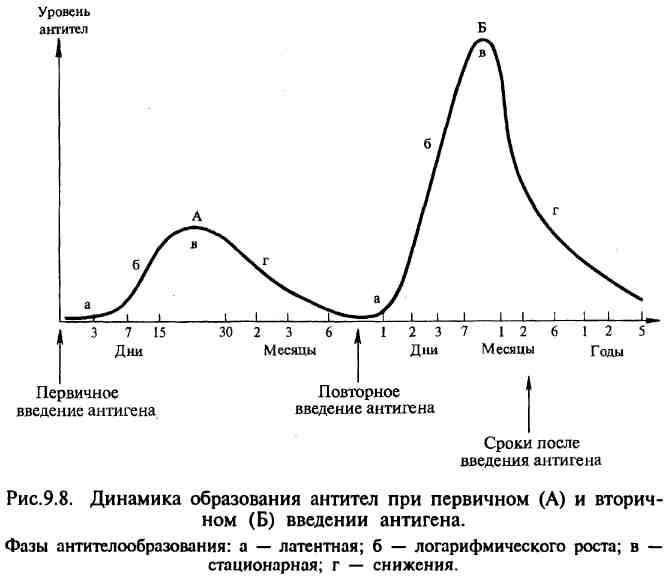 В латентной фазе происходят переработка и представление антигена иммунокомпетентным клеткам, размножение клона клеток, специализированного на выработку антител к данному антигену, начинается синтез ан тител. В этот период антитела в крови не обнаруживаются. Во время логарифмической фазы синтезированные антитела высво бождаются из плазмоцитов и поступают в лимфу и кровь. В ста ционарной фазе количество антител достигает максимума и ста билизируется, затем наступает фаза снижения уровня антител. При первичном введении антигена (первичный иммунный от вет) латентная фаза составляет 3—5 сут, логарифмическая — 7— 15 сут, стационарная — 15—30 сут и фаза снижения — 1—6 мес и более. Особенностью первичного иммунного ответа является то, что первоначально синтезируется IgM, а затем IgG. В отличие от первичного иммунного ответа при вторичном введении антигена (вторичный иммунный ответ) латентный период укорочен до нескольких часов или 1—2 сут, логарифми ческая фаза характеризуется быстрым нарастанием и значитель но более высоким уровнем антител, который в последующих фазах длительно удерживается и медленно, иногда в течение не скольких лет, снижается. При вторичном иммунном ответе в отличие от первичного синтезируются главным образом IgG. Такое различие динамики антителообразования при первич ном и вторичном иммунном ответе объясняется тем, что после первичного введения антигена в иммунной системе формирует ся клон лимфоцитов, несущих иммунологическую память о данном антигене. После повторной встречи с этим же антиге ном клон лимфоцитов с иммунологической памятью быстро раз множается и интенсивно включает процесс антителогенеза. Очень быстрое и энергичное антителообразование при повтор ной встрече с антигеном используется в практических целях при необходимости получения высоких титров антител при произ водстве диагностических и лечебных сывороток от иммунизиро ванных животных, а также для экстренного создания иммуни тета при вакцинации. 40. Классификация гиперчувствительности по Джейлу и Кумбсу. Стадии аллергической реакции. Изучение молекулярных механизмов аллер гии привело к созданию Джеллом и Кумбсом в 1968 г. новой классификации. В соответствии с ней различают четыре основных типа аллер гии: анафилактический (I тип), цитотоксический (II тип), иммунокомплексный (III тип) и опосредованный клетками (IV тип). Первые три типа относятся к ГНТ, четвертый — к ГЗТ. Ведущая роль в запуске ГНТ играют антитела (IgE, G и М), а ГЗТ — лимфоидно-макрофагальная реакция. Аллергическая реакция I типа связана с биологическими эффектами IgE и G4, на званных реагинами, которые обладают цитофильностью — сродством к тучным клеткам и базофилам. Эти клетки несут на поверхности высокоаффинный FcR, связывающий IgE и G4 и использующий их как ко-рецепторный фактор специфического взаимодействия с эпитопом аллергена. Связывание аллергена с рецепторным комплексом вызывает дегрануляцию базофила и тучной клетки — залповый выброс биологически активных соединений (гистамин, гепарин и др.), содержащихся в гранулах, в межклеточное пространство. В результате развиваются бронхоспазм, вазодилатация, отек и прочие симптомы, характерные для анафилаксии. Вырабатываемые цитокины стимулируют клеточное звено иммунитета: образование Т2-хелпера и эозинофилогенез. Цитотоксические антитела (IgG, IgM), на правленные против поверхностных структур (антигенов) соматических клеток макроорга низма, связываются с клеточными мембра нами клеток-мишеней и запускают различ ные механизмы антителозависимой цитотоксичности (аллергическая реакция II типа). Массивный цитолиз сопровождается соот ветствующими клиническими проявлениями. Классическим примером является гемолити ческая болезнь в результате резус-конфликта или переливания иногруппной крови. Цитотоксическим действием обладают так же комплексы атиген—антитело, образующи еся в организме пациента в большом количес тве после введения массивной дозы антигена (аллергическая реакция III типа). В связи с кумулятивным эффектом клини ческая симптоматика аллергической реакции III типа имеет отсроченную манифестацию, иногда на срок более 7 суток. Тем не менее этот тип реакции относят к ГНТ. Реакция может проявляться как одно из осложнений от при менения иммунных гетерологичных сыворо ток с лечебно-профилактической целью («сы вороточная болезнь»), а также при вдыхании белковой пыли («легкое фермера»). Лабораторная диагностика аллергии при аллергических реакциях I типа основана на выявлении суммарных и специфических реа гинов (IgE, IgG4) в сыворотке крови пациен та. При аллергических реакциях II типа в сы воротке крови определяют цитотоксические антитела (антиэритроцитарные, антилейко цитарные, антитромбоцитарные и др.). При аллергических реакциях III типа в сыворотке крови выявляют иммунные комплексы. Для обнаружения аллергических реакций IV ти па применяют кожно-аллергические пробы, которые широко используют в диагностике некоторых инфекционных и паразитарных заболеваний и микозов (туберкулез, лепра, бруцеллез, туляремия и др.).
|