шиза. 1. Общие свойства возбудимых тканей. Возбудимость
 Скачать 354.5 Kb. Скачать 354.5 Kb.
|

|
1. Общие свойства возбудимых тканей. Возбудимость – способность ткани отвечать на раздражение изменением ряда своих свойств. Показатель возбудимости – порог раздражения. Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани. Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения возбуждения. Проводимость напрямую зависит от возбудимости ткани: чем выше возбудимость, тем выше проводимость, так как быстрее возбуждается расположенный рядом участок ткани. Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения. При этом в ходе ответной реакции ткань перестает воспринимать раздражитель. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель). Показатель рефрактерности (рефрактерный период) - время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани Лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность). Для мышечной ткани характерна также сократимость. Сократимость – способность мышцы отвечать сокращением на раздражение. 2.Классификация раздражителей Раздражитель – фактор, способный вызвать ответную реакцию возбудимых тканей. 1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах); 2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т. п.), физико-химические (осмотические – кристаллик хлорида натрия). По своейприродераздражители бывают:
Побиологическому соответствию, то есть насколько раздражитель соответствует данной ткани. 1) адекватные – раздражители, которые соответствуют данной ткани. Например, для сетчатки глаза свет – все остальные раздражители не соответствуют сетчатке, для мышечной ткани – нервный импульс и т.д.; 2) неадекватные – раздражители, которые не соответствуют данной ткани. Для сетчатки глаза все раздражители кроме светового будут неадекватные, а для мышечной ткани все раздражители, кроме нервного импульса. Посиле: 1) подпороговые раздражители – это сила раздражителя при которой не возникает ответная реакция; 2) пороговый раздражитель – это минимальная сила, которая вызывает ответную реакцию при бесконечном времени действия. Эту силу еще называют реобазой – она единственная для каждой ткани; 3) надпороговые, или субмаксимальные; 4)максимальный раздражитель– это минимальная сила при которой возникает максимальная ответная реакция ткани; 5) сверхмаксимальные раздражители – при этих раздражителях реакция ткани либо максимальная, либо уменьшается, либо временно исчезает. Таким образом, для каждой ткани существует один пороговый раздражитель, один максимальный и множество подпороговых, надпороговых и сверхмаксимальных. 3. Физиология клеточных мембран. Механизмы трансмембранного транспорта. - Пограничная функция. Мембрана отграничивает цитоплазму от межклеточной жидкости, а большинство внутриклеточных структур: митохондрии, ядро, эндоплазматическую сеть – от цитоплазмы. - Биотрансформирующая функция. Любое вещество, проходя через мембрану, вступает с ней в сложное взаимодействие и претерпевает ряд биохимических превращений. В результате биотрансформации лекарственное вещество, как правило, переходит в форму, легко усвояемую клеткой. - Транспортная функция. Перенос веществ через биологические мембраны связан с процессами метаболизма, поддержанием постоянства внутренней среды клетки, возбуждением и проведением нервного импульса. Существует два основных типа переноса: пассивный (фильтрация, диффузия, облегченная диффузия, осмос) и активный (работа мембранных белковых «насосов») Пассивный транспорт. Фильтрация осуществляется через мембранные белковые каналы – поры, зависит от разности давлений снаружи и внутри клетки и проницаемости мембраны для жидкости и низкомолекулярных веществ. Диаметр пор чрезвычайно мал, поэтому фильтруются только низкомолекулярные вещества, вода и некоторые ионы. Диффузия - пассивное передвижение молекул или ионов по градиенту концентрации (из области высокой концентрации в область низкой). Осмос представляет собой частный случай диффузии растворителя через полупроницаемую мембрану, не пропускающую растворенные вещества. Пассивный транспорт не требует затрат энергии. Активный транспорт. Это универсальный для всех видов мембран перенос веществ против концентрационных или электрохимических градиентов (из области низкой концентрации в область высокой). При помощи активного транспорта переносятся гидрофильные полимерные молекулы, неорганические ионы (Na, Ca, K) , водород, сахара, аминокислоты, витамины, гормоны и лекарственные вещества. Активный транспорт осуществляется с обязательной затратой энергии, образующейся при расщеплении (окислительное фосфорилирование) аденозинтрифосфорной кислоты (АТФ). Разновидностью активного транспорта, связанной с деятельностью самой клетки, является микровезикулярный транспорт (пиноцитоз, экзоцитоз и фагоцитоз). При пиноцитозе происходит активное поглощение клеткой жидкости из окружающей среды с формированием пузырьков и последующим переносом их через цитоплазму. Процесс слияния пузырьков с мембраной клетки и выделение клеткой вещества в виде секреторных гранул или вакуолей называется экзоцитозом. Явление фагоцитоза заключается в способности клеток активно захватывать и поглощать микроорганизмы, разрушенные клетки и инородные частицы. - Рецепторная функция. Биологические мембраны имеют большое количество рецепторов – участков, молекулярная структура которых характеризуется избирательным сродством к определенным физиологически активным веществам: гормонам, медиаторам, антигенам. - Образование межклеточных контактов. - Генерация биоэлектрических потенциалов. В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала. 4. Ионные механизмы мембранного потенциала покоя О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет видимого функционального отправления ткани. Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией; 2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концентрации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил: 1) силы диффузии; 2) силы электростатического взаимодействия. Значение электрохимического равновесия: 1) поддержание ионной асимметрии; 2) поддержание величины мембранного потенциала на постоянном уровне. В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентрационно-электрохимическим. Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает: 1) высокую концентрацию ионов К внутри клетки, т. е. постоянную величину потенциала покоя; 2) низкую концентрацию ионов Na внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия; 3) стабильный концетрационный градиент ионов Na, способствуя транспорту аминокислот и сахаров. 5. Мембранный потенциал действия: фазы, ионные механизмы. Потенциал действия– это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия. Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа+-канала (1). Ионы Na+устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала (2). При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время (3). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы (2) и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциалпринимает первоначальное значение (3), и даже превышает на короткое времяпотенциал покоя (4). После этогонервная клетка вновь становится возбудимой. За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоихионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткойновых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращениемембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку. Восходящая ветвь графика:
Нисходящая ветвь графика:
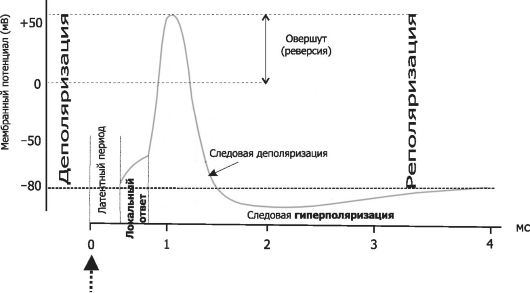 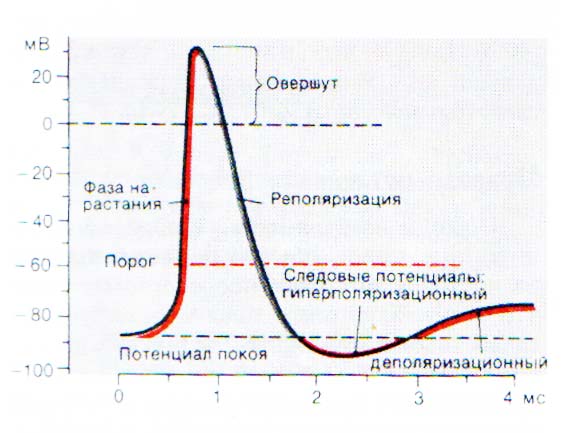 6. Классификация нервных волокон. А – нервные волокна с самой толстой миелиновой оболочкой. Наиболее высокая скорость передачи нервного импульса. В – миелиновая оболочка тоньше, скорость проведения возбуждения ниже С – безмиелиновые волокна с относительно низкой скоростью передачи импульса.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||