шиза. 1. Общие свойства возбудимых тканей. Возбудимость
 Скачать 354.5 Kb. Скачать 354.5 Kb.
|

|
Типы нервных волокон, их свойства и функциональное назначение
Нервные волокна всех групп обладают общими свойствами: 7. Проведение возбуждения по немиелинизированным волокнам В безмиелиновых волокнах нервный импульс распространяется волнообразно, последовательно возбуждаются небольшие участки размером в несколько микрометров. Возбуждение распространяется гораздо медленнее, чем по миелиновым волокнам. В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление. Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна. При действии раздражителя пороговой силы на мембрану безмиелинового волокна изменяется ее проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны (внутренняя становиться заряженной положительно, а наружная отрицательно). Это ведет к возникновению круговых токов (заряженных частиц) от «+» к «–» на протяжении всего волокна. Особенности распространения возбуждения по безмиелиновым волокнам: 1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением. 2. Возбуждение распространяется с небольшой скоростью. 3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна). По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров. 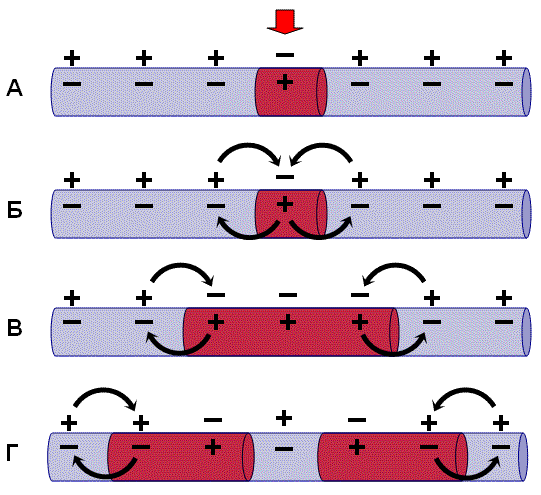 8. Проведение возбуждения по миелинизированным волокнам Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированном волокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности. Особенности распространения возбуждения по миелиновым волокнам: 1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением. 2. Возбуждение распространяется с большой скоростью. 3. Возбуждение распространяется без декримента. По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции. В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье. При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому. 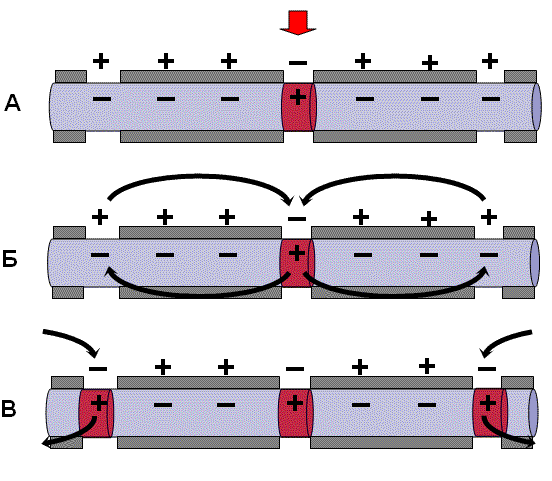 9. Законы проведения возбуждения по нервному волокну • Закон анатомической и физиологической непрерывности –возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности. • Закон двустороннего проведения возбуждения– возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно). • Закон изолированного проведения– возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна. 10. Связь возбудимости с фазами потенциала действия Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением. В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности. В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности. Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости. В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит. 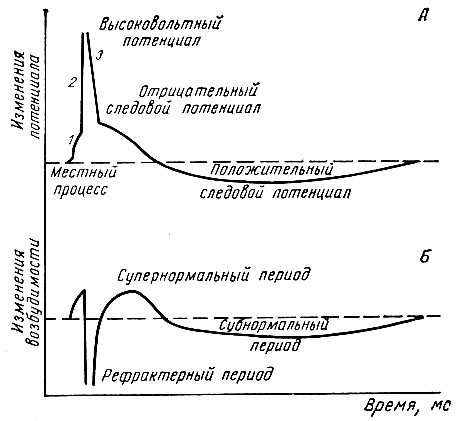 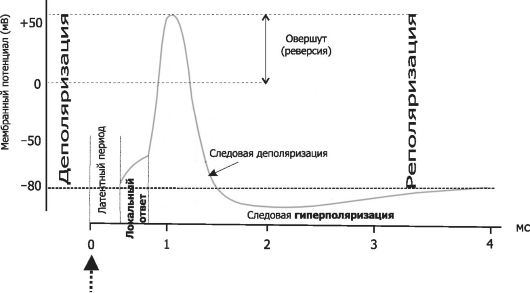 11. Полярный закон раздражения. Физиологический электротон. Закон полярного действия постоянного тока(закон Пфлюгера): при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом. Прохождение постоянного электрического тока через нервное или мышечное волокно вызывает изменение мембранного потенциала покоя. Так, в области приложения к возбудимой ткани катода положительный потенциал на наружной стороне мембраны уменьшается, возникает деполяризация, которая быстро достигает критического уровня и вызывает возбуждение. В области же приложения анода положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает. Но при этом под анодом критический уровень деполяризации смещается к уровню потенциала покоя. Поэтому при размыкании цепи тока гиперполяризация на мембране исчезает и потенциал покоя, возвращаясь к исходной величине, достигает смещенного критического уровнями возникает возбуждение. Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом и соседних с ним участках понижается вследствие деполяризации мембраны - возбудимость повышается. В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости). Повышение возбудимости под катодом называется катэлектротоном, а снижение возбудимости под анодом - анэлектротоном. При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением, развивается так называемая катодическая депрессия. Первоначальное же снижение возбудимости под анодом сменяется ее повышением - анодная экзальтация. При этом в области приложения катода происходит инактивация натриевых каналов, а в области действия анода происходит снижение калиевой проницаемости и ослабление исходной инактивации натриевой проницаемости. 12. Моторные единицы. Механизм сокращения и расслабления мышечных волокон. Анатомической и функциональной единицей скелетных мышц является нейромоторная единица - двигательный нейрон и иннервируемая им группа мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы небольших мышц содержат малое число мышечных волокон, а ДЕ крупных мышц туловища и конечностей — до нескольких тысяч. Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга. Поперечно-полосатая скелетная мышца состоит из длинных волокон – миофибрилл, внутри которых располагаются нити сократительных белков – актина и миозина. Нити миозина толстые и не смещаются, из актина образуются тонкие нити, способные к смещению. Нити актина покрыты слоем белка тропонина, препятствующего их взаимодействию с миозином. Нити сократительных белков окружены цитоплазмой (саркоплазмой). Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий. При возбуждении мышцы открываются многочисленные каналы в мембране саркоплазматической сети, через которые в саркоплазму выходят ионы кальция. Они взаимодействуют с белком тропонином, освобождая от него нити актина. Кроме того, кальций стимулирует распад молекул АТФ, высвобождая большое количество энергии, за счет которой актин взаимодействует с миозином – нити актина как бы вдвигаются в промежутки между нитями миозина, и все мышечное волокно сокращается. Для расслабления мышцы тоже требуется энергия в виде молекул АТФ. За счет этой энергии идет работа кальциевого насоса, удаляющего ионы кальция из саркоплазмы. В результате освободившиеся молекулы тропонина блокируют актин, препятствуя его взаимодействию с миозином. Нити снова расходятся, мышечное волокно расслабляется. Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается. 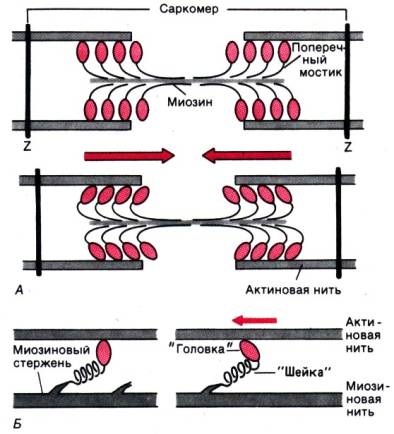 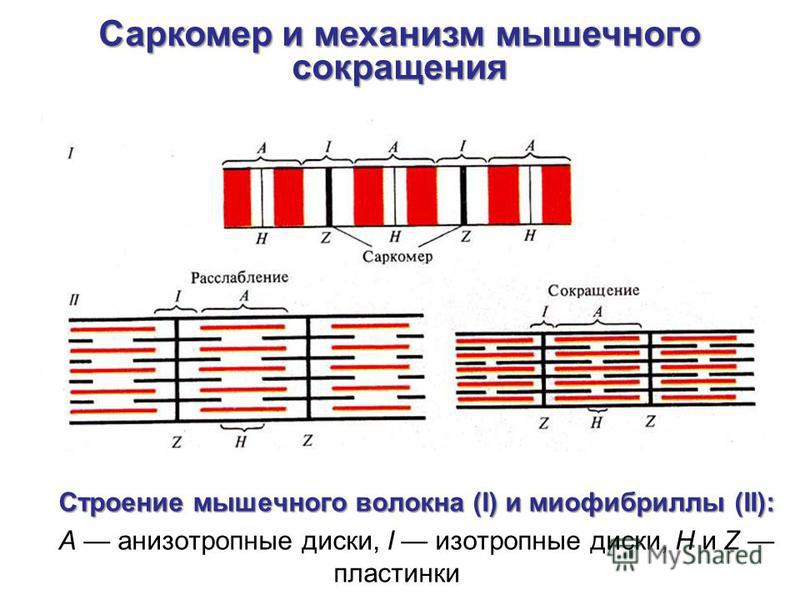 13. Типы скелетных мышечных волокон Красные мышечные волокна (волокна 1 типа) содержат большое количество митохондрий с высокой активностью окислительных ферментов. Сила их сокращений сравнительно невелика, а скорость потребления энергии такова, что им вполне хватает аэробного метаболизма. Они участвуют в движениях, не требующих значительных усилий, - например, в поддержании позы. Присутствует миоглобин. Белым мышечным волокнам (волокнам 2 типа) присуща высокая активность ферментов гликолиза, значительная сила сокращения и такая высокая скорость потребления энергии, для которой уже не хватает аэробного метаболизма. Поэтому двигательные единицы, состоящие из белых волокон, обеспечивают быстрые, но кратковременные движения, требующие рывковых усилий. 14. Одиночное мышечное сокращение. Его фазы. Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений. Одиночное мышечное сокращение.При раздражении мышцы одиночным импульсом возникает одиночное мышечное сокращение, в котором выделяют три фазы:
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока. 15. Связь возбудимости с фазами одиночного мышечного сокращения. В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий. • В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану. • Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В). • Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г). • К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д). Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения. Раздражение -- возникновение ПД -- проведение его вдоль клеточной мембраны и вглубь волокна по трубочкам Т-систем -деполяризация мембраны саркоплазматического ретикулюма -- освобождение Са++ из триад и диффузия его к миофибриллам -- взаимодействие Са++ с тропонином и выделение энергии АТФ -- взаимодействие (скольжение) актиновых и миозиновых нитей -- сокращение мышцы -- понижение концентрации Са++ в межфибриллярном пространстве из-за работы Са-насоса -- расслабление мышцы. 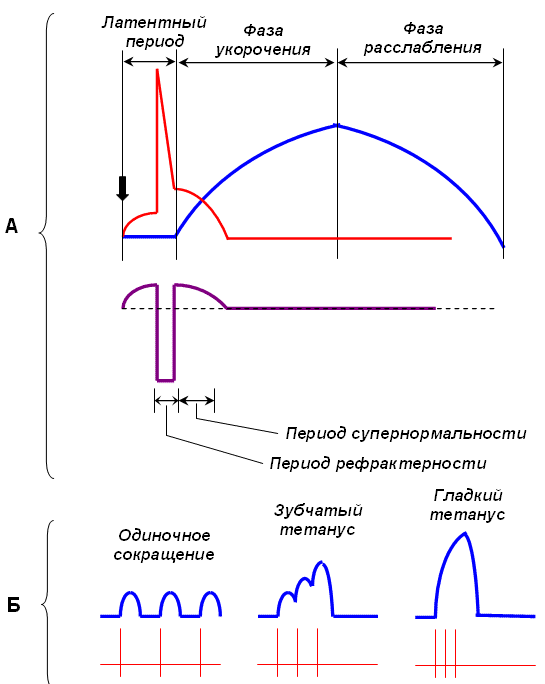 |