1. Понятие о клеточном цикле. Фазы клеточного цикла их характеристика и продолжительность
 Скачать 191.48 Kb. Скачать 191.48 Kb.
|

|
10.Нарушения мейоза. при делении соматических клеток могут возникнуть нарушения, связанные с повреждением хромосом, митотического аппарата, цитоплазмы. Задержка митоза в профазе, нарушение спирализации и диспирализации, раннего разделения хроматид. Эти нарушения возникают под действием отдельных хим. веществ, радиации, вирусных инфекций. Основные патологии мейоза – не расхождение хромосом: первичная, вторичная, третичная. Первичная – у особей с нормальным кариотипом: в анафазе I нарушение разделения бивалентов и обе хромосомы из пары гомологов переходят в одну клетку., что приводит к избытку хромосом в данной клетке и недостатка в другой. Полиплодия (от греческих слов polyploos — многократный и eidos — вид) — наследственное изменение, заключающееся в кратком увеличении числа наборов хромосом в клетках. Дети всегда похожи на обоих родителей. Происходит это потому, что в каждой их клетке находятся два набора хромосом, два комплекта генов — один материнский и один отцовский. Такой двойной, или диплоидный (от греческих слов diploos — двойной и eidos — вид), набор хромосом типичен для живой природы. Он достаточен для преемственности поколений. Но в некоторых тканях диплоидных организмов в процессе их развития появляются клетки, в которых 4, 8 или гораздо больше наборов хромосом. Такие клетки называют полиплоидными, а сам процесс — соматической полиплоидией ,Многие виды растений полиплоидные. Вероятно, так же эволюционировали и некоторые животные. Примером могут служить некоторые черви, насекомые, рыбы и др. Человек давно использует полиплоидию для выведения высокопродуктивных сортов сельскохозяйственных растений. Вторичная – нерасхождение возникает в гаметах у особей с избытком одной хромосомы в кариотипе (образуются биваленты и униваленты). Из всех синдромов, возникающих вследствие нерасхождения аутосом, наиболее распространенной патологией является болезнь Дауна (95 % от числа всех трисомий по аутосомам). При болезни Дауна наблюдается трисомия 21-й хромосомы. Клиническими признаками болезни являются низкий рост, широкое круглое лицо, близко расположенные глаза с узкими глазными щелями, полуоткрытый рот. Для болезни характерны также идиотия и дефекты сердечно-сосудистой системы (пороки сердца и крупных сосудов). При синдроме Эдвардса, характеризующемся трисомией по 18-й хромосоме, имеются множественные физические пороки развития: общая гипотрофия новорожденного, задержка психомоторного развития, крипторхизм, порок сердца, грыжи и многие другие. Синдром Патау, трисомия по 13-й хромосоме, характеризуется микроцефалией, полидактилией, наличием расщелины верхней губы и неба. Частичные трисомии и частичные моносомии выявляются при анализе причин врожденных дефектов развития новорожденных детей. Несбалансированность по генам каждой из хромосом проявляется у новорожденных в виде специфических признаков. Так, например, частичная моносомия короткого плеча 5-й хромосомы дает патологию, описанную как синдром "кошачьего крика" при котором имеются аномалии развития нижней челюсти и гортани, что сопровождается характерным изменением голоса, а также микроцефалия, пороки сердца, четырехпалость и др. При нерасхождении половых хромосом формируется группа синдромов, для которых с клинической точки зрения наиболее характерны интеллектуальное и половое недоразвитие наряду с физическими дефектами. Так, при синдроме Тернера — Шерешевского у пациента женского пола (генотип ОХ) обнаруживаются отставание в развитии (низкий рост), половой инфантилизм, бесплодие, иногда умственная отсталость, пороки сердца и др. У женщины при трисомий X (генотип XXX) имеются умственная отсталость и нарушения физического развития. При синдроме Клайнфелтера (генотип XXY) или сверх Клайнфелтера — (XXXY) наблюдается высокий рост с непропорционально длинными конечностями, гипоплазия яичек, недоразвитие вторичных половых признаков, бесплодие, склонность к асоциальному поведению. Третичная – имеет структурные перестройки хромосом. Структурные перестройки хромосом, какого бы вида они ни были, вызывают нарушения развития организма вследствие или недостатка части материала по данной хромосоме (частичная моносомия) или его избытка (частичная трисомия). Как пример можно привести Х-полисомию при отсутствии У-хромосомы. Такие организмы имеют хромосомный набор 47,XXX и хотя внешне женщины выглядят нормально и они плодовиты, но у них отмечается умственная отсталость. При синдроме Клайнфельтера (47,ХХУ) мужчина обладает некоторыми вторичными женскими подовыми признаками, бесплоден, яички слабо развиты, волос на лице мало, иногда развиваются молочные железы, обычно низкий уровень умственного развития. При хромосомном наборе 47.ХУУ мужчины имеют высокий рост, различный уровень умственного развития, иногда обладают психопатическими чертами или проявляют склонность к мелким правонарушениям. 11.Мейоз, цитологическая и цитогенетическая характеристика, биологическое значения. Мейоз – это особое деление соматических клеток половых желез, которое приводит к образованию гамет. Мейоз состоит из двух делений – мейоз I и мейоз II. Каждое деление имеет четыре фазы: профаза I и профаза II, метафаза I и метафаза II, анафаза I и анафаза II, телофаза I и телофаза II Самой сложной является профаза мейоза I. Она имеет 5 стадий: 1- лептотена: хроматин спирализуется, образуются тонкие хроматиновые нити, которые начинают движение друг к другу центромерными участками; генетический материал – 2n2chr4c. 2 – зиготена: начинается конъюгация коротких, толстых хроматиновых нитей (хромосом), которые соединяются по всей длине; генетическая информация не изменяется – 2n 2chr4c. 3 – пахитена: гомологичные хромосомы плотно соединяются по всей длине; образуемые фигуры называют бивалентыхромосом или тетрады хроматид; генетический материал можно записать так – 1n biv4chr4с; к концу стадии в области центромер начинают действовать силы отталкивания и происходит кроссинговер – обмен участками гомологичных хромосом; 4 – диплотена: продолжают действовать силы отталкивания, но хромосомы остаются соединенным в области хиазм (перекрестов); содержание генетического материала сохраняется – 1n biv 4chr4с; 5 – диакинез: заканчивается спирализация хромосом, исчезают ядерная оболочка и ядрышко; биваленты хромосом, соединенные своими концами, выходят в цитоплазму и движутся к центру клетки; нити веретена деления прикрепляются к центромерам хромосом; 1n biv 4chr4c. В метафазе мейоза I биваленты располагаются по экватору клетки; четко видны отдельные хромосомы; генетический материал – 1n biv 4chr4с. Анафаза I: биваленты распадаются на отдельные гомологичные хромосомы, которые расходятся к полюсам клетки; каждая хромосома содержит 2 хроматиды; содержание генетического материала на каждом полюсе клетки – 1n2chr2с; произошла редукция (уменьшение) числа хромосом – из диплоидного набор хромосом стал гаплоидным. Поэтому первое деление мейоза называется редукционным. В телофазу мейоза I происходит цитокинез, и образуются две дочерние гаплоидные клетки – 1n2chr2c; в отличие от митоза в этой фазе не происходит деспирализации хромосом. После мейоза I наступает интеркинез – короткий промежуток между двумя делениями и начинается мейоз II. Репликация ДНК не происходит. Второе деление мейоза не отличается от митоза, но в профазе II не происходит спирализации хромосом (1n2chr2c), а в анафазе II к полюсам клетки отходят хроматиды (дочерние хромосомы). Каждая дочерняя клетка получает набор генетической информации1n1chr1с. Гаплоидный набор хромосом сохраняется. Второе деление мейоза называется эквационным, или уравнительным. Из одной материнской диплоидной клетки образуются 4 клетки (гаметы) с гаплоидным набором хромосом. Значение мейоза: - это механизм образования гамет; - поддерживает постоянство числа хромосом при половом размножении; - обеспечивает комбинативную изменчивость в результате кроссинговера, независимого расхождения хроматид и хромосом, при образовании гамет. 12. Законы Менделя: Закономерности наследования при могоногибридном скрещивании и их цитологические основы. Менделериющие признаки человека. Первый закон Менделя (закон единообразия): В первом поколении все гибриды одинаковы, похожи на одного из родителей. Второй закон Менделя (закон расщепления): При скрещивание гетерозиготных гибридов первого поколения происходит расщепление признаков в соотношении 3:1. Третий закон Менделя (закон независимого наследования признаков): При дигибридном скрещивании расщепление по каждой паре признаков идет независимо от других признаков. Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. При этом скрещиваемые предки являются гетерозиготными по положению хромосомы в аллели. Моногибридное наследование представляет собой пример наследования единственного признака (гена), различные формы которого называют аллелями. Например, при моногибридном скрещивании между двумя чистыми линиями растений, гомозиготных по соответствующим признакам — одного с жёлтыми семенами (доминантный признак), а другого с зелёными семенами (рецессивный признак), можно ожидать, что первое поколение будет только с жёлтыми семенами, потому что аллель жёлтых семян доминирует над аллелью зелёных. При моногибридном скрещивании сравнивают только один характерный признак. Источник: Википедия Грегор Мендель (1822 - 1884) — выдающийся чешский ученый. Основоположник генетики. Впервые обнаружил существование наследственных факторов, впоследствии названных генами. Грегор Мендель проводил опыты с горохом. Серди большого количества сортов он выбрал для первого эксперимента два, отличающихся по одному признаку. Семена одного сорта гороха были желтые, а другого — зеленые. Известно, что горох, как правило, размножается путем самоопыления и поэтому в пределах сорта нет изменчивости по окраске семян. Используя это свойство гороха, Г. Мендель произвел искусственное опыление, скрестив сорта, отличающиеся цветом семян (желтым и зеленым). Независимо от того, к какому сорту принадлежали материнские растения, гибридные семена оказались только желтыми. Цитологические основы закономерностей наследования при моногибридном скрещивании Изобразим моногибридное скрещивание в виде схемы. Символ 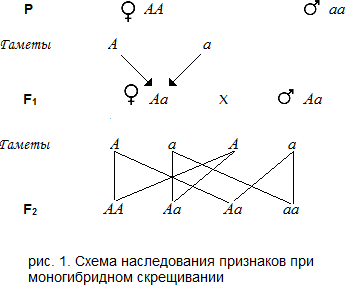 Из рисунка видно, что в каждой гамете родительских особей будет по одному гену: в одном случае A, в другом — а. Таким образом, в первом поколении все соматические клетки будут гетерозиготными — Aa. В свою очередь, гибриды первого поколения с равной вероятностью могут образовывать гаметы A или a. Случайные комбинации этих гамет при половом процессе могут дать следующие варианты: AA, Aa, aA, aa. Первые три растения содержащие ген A, по правилу доминирования будут иметь желтые горошины, а четвертое — рецессивная гомозигота aa — будет иметь зеленые горошины. Из рисунка видно, что в каждой гамете родительских особей будет по одному гену: в одном случае A, в другом — а. Таким образом, в первом поколении все соматические клетки будут гетерозиготными — Aa. В свою очередь, гибриды первого поколения с равной вероятностью могут образовывать гаметы A или a. Случайные комбинации этих гамет при половом процессе могут дать следующие варианты: AA, Aa, aA, aa. Первые три растения содержащие ген A, по правилу доминирования будут иметь желтые горошины, а четвертое — рецессивная гомозигота aa — будет иметь зеленые горошины.Законы Менделя справедливы для моногенных признаков, которые также называют менделирующими. Чаще всего их проявления носят качественный альтернативный характер: карий и голубой цвет глаз, нормальная свертываемость крови или гемофилия. В формировании других признаков, таких как рост, вес, характер телосложения или тип поведения, могут участвовать десятки или даже сотни генов. Степень выраженности подобных признаков у отдельных особей часто может быть измерена количественно, и потому такие признаки называют количественными.  13.. Законы Менделя: Закономерности наследования при дигибридном скрещивании и их цитологические основы. Менделериющие признаки человека. Первый закон Менделя (закон единообразия): В первом поколении все гибриды одинаковы, похожи на одного из родителей. Второй закон Менделя (закон расщепления): При скрещивание гетерозиготных гибридов первого поколения происходит расщепление признаков в соотношении 3:1. Третий закон Менделя (закон независимого наследования признаков): При дигибридном скрещивании расщепление по каждой паре признаков идет независимо от других признаков. Дигибридное скрещивание. Закон независимого наследования признаков. Скрещивание, при котором родительские формы отличаются по двум парам альтернативных признаков, называется дигибридным. Если родители отличаются по многим парам альтернативных признаков, скрещивание называется полигибридным. В опытах Г Мендель скрещивал гомозиготные растения гороха, различающиеся одновременно по двум парам признаков, например по окраске и форме поверхности семян. Одни родительские растения имели желтые гладкие семена, другие — зеленые морщинистые. В первом поколении наблюдалось единообразие — все гибриды имели желтые гладкие семена. Значит, желтая окраска полностью доминирует над зеленой, а гладкая поверхность — над морщинистой. Путем самоопыления гибридов первого поколения было получено второе поколение, в котором согласно закону расщепления проявились не только доминантные, но и рецессивные признаки. При этом наблюдались все возможные сочетания признаков семян: желтые гладкие, желтые морщинистые, зеленые гладкие и зеленые морщинистые в соотношении 9:3:3: 1. Таким образом, было получено потомство четырех фенотипических классов: доминантные по обоим признакам — —; доминантные по первому и рецессивные по второму признаку — —, рецессивные по первому и доминантные по второму — у|-, рецессивные по обоим признакам — часть. Это говорит о том, что при дигибридиом (а также полигибридном) скрещивании расщепление по каждой паре альтернативных признаков происходит н е -зависимо от других. Значит, дигибридное скрещивание, по сути, представляет собой два независимо идущих моногибридных скрещивания. Эта закономерность, установленная Г. Менделем, впоследствии была названа законом независимого наследования признаков или третьим законом Менделя. Его можно сформулировать следующим образом: при скрещивании особей, отличающихся по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо и комбинируются во всех возможных сочетаниях. 14.Взаимодействие аллельных генов. Примеры из жив и раст мира .Аллельные гены - это гены, которые занимают одинаковые «места» (или локусы) в хромосомах Доминирование — это такое взаимодействие аллельных генов, при котором проявление одного из аллелей (А) не зависит от присутствия в генотипе другого аллеля (А') и гетерозиготы АА' фенотипически не отличаются от гомозигот по этому аллелю (АА). например, когда один из аллелей гена А («дикий») способен обеспечить формирование определенного варианта признака (синтез пептида с определенными свойствами), а другой А'—не обладает такой способностью. Наличие в генотипе АА' единственного нормального аллеля А приводит к формированию нормального признака. Этот аллель выступает как доминантный в данном гетерозиготном генотипе. Присутствие другого аллеля (А') фенотипически не проявляется, поэтому его называют рецессивным. Примером доминирования 1го из алледей в гетерозиготном генотипе может служить определение групповой принадлежности крови у человека по системе АВ0. Генотипы, содержащие аллель IA либо в гомозиготном состоянии, либо в сочетании с аллелем I0 (IAIA или IAI0), определяют развитие у человека второй группы крови (группа крови А). Такая же ситуация наблюдается и в отношении аллеля IB, обусловливающего формирование третьей, или В-группы крови. Следовательно, аллели IA и IB выступают как доминантные по отношению к аллелю I0, формирующему в гомозиготном состоянии I0I0 первую, или 0-группу крови. Неполное доминирование наблюдается, когда фенотип гетерозигот BB' отличается от фенотипа гомозигот по обоим аллелям (BB или B'B') промежуточным проявлением признака. Это объясняется тем, что аллель, способный сформировать нормальный признак, находясь в двойной дозе у гомозиготы BB, проявляется сильнее, чем в единственной дозе у гетерозиготы BB'. Указанные генотипы отличаются экспрессивностью, т.е. степенью выраженности признака. Демонстрацией такого типа взаимодействия генов могут быть многочисленные наследственные заболевания у человека, проявляющиеся клинически у гетерозигот по мутантным аллелям, а у гомозигот заканчивающиеся смертью. Иногда гетерозиготы имеют почти нормальный фенотип, а гомозиготы характеризуются пониженной жизнеспособностью. Так, гомозиготы по аллелюсерповидноклеточности эритроцитов в связи с развитием у них тяжелой формы анемии и других фенотипических проявлений обычно не переживают детский возраст. Напротив, гетерозиготы — это как правило нормальные люди. Вместе с тем, они все-таки испытывают кислородную недостаточность в большей степени в сравнении с гомозиготами по аллелю дикого типа, в частности при подъеме на высоту Кодоминированиепредставляет собой такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате этого формируется некий промежуточный вариант признака, новый по сравнению с вариантами, определяемыми каждым аллелем самостоятельно. Н:формирование IV, или АВ-группы, крови у человека, гетерозиготного по аллелям IA и IB, которые по отдельности детерминируют образование II и III групп крови. Межаллельнаякомплементация относится к достаточно редко встречаемым способам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям гена D(D'D"). Допустим, что ген D отвечает за синтез какого-то белка, который имеет четвертичную структуру, состоящую из нескольких одинаковых пептидных цепей. Мутантный аллель D' определяет синтез измененного пептида D', a мутантный аллель D" приводит к синтезу другой, но тоже измененной структуры пептида D". Можно представить ситуацию, когда взаимодействие таких измененных пептидов (D' и D") при формировании четвертичной структуры, как бы взаимно компенсируя эти изменения, обеспечивает образование белка с нормальными свойствами. В то же время отдельно взаимодействующие пептиды D' или D" формируют аномальные белки. Таким образом, с определенной вероятностью у гетерозигот D'D" в результате межаллельнойкомплементации может образовываться нормальный признак в виде белка с нормальными свойствами. ТО, даже процесс формирования элементарного признака — синтез полипептида с определенной последовательностью аминокислот — зависит, как правило, от взаимодействия по меньшей мере двух аллельных генов, и конечный результат определяется конкретным сочетанием их в генотипе. 15.Взаимодействие неаллельных генов. Примеры из жив и раст мира Неалле́льныеге́ны — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов.Большинство количественных признаков организмов определяется полигенами, т.е. системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов называют полимерным. Оно сводится чаще всего к суммированию действия сходных аллелей этих генов, определяющих формирование одинакового варианта признака. По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня отложения в клетках пигмента меланина. В геноме человека имеется четыре гена, отвечающих за этот признак. В генотипе все они представлены в двойной дозе. В связи с тем что эти гены отвечают за развитие одного и того же признака, их обозначают одной и той же буквой, но с разными символами, чтобы подчеркнуть их неаллельность — P1P2P3P4. Наличие в генотипе восьми доминантных аллелей в системе полигенов, определяющих цвет кожи, обусловливает максимальную ее пигментацию, наблюдаемую у африканских негров (P1P1P2P2P3P3P4P4 Полимерное взаимодействие генов лежит в основе определения главным образом количественных признаков (рост, масса организма, возможно, интеллект). . Такое взаим-е неаллельных генов, при кот-м они взаимно дополняют друг друга, называют комплементарным взаимодействием.Например процесс формирования половой принадлежности организма у человека. Развитие признаков пола у человека, так же как у большинства животных, определяется в первую очередь сочетанием гетерохромосом в его кариотипе. Наличие Х- и Y-хромосом обусловливает возможность формирования мужского пола, а двух Х-хромосом —женского. Однако установлено, что для развития организма мужского пола не достаточно присутствия одного лишь Y-сцепленного гена, который определяет дифференцировку половых желез по мужскому типу и синтез ими гормона тестостерона. Для этого необходим также продукт другого гена — белок-рецептор, обеспечивающий проникновение гормона в клетки тканей-мишеней. За синтез такого белка отвечает особый ген, расположенный в Х-хромосоме. Его мутация, нарушающая образование нормального белка-рецептора, делает ткани-мишени невосприимчивыми к гормону, направляющему их развитие по мужскому типу. Не использовав такую возможность на определенном этапе онтогенеза, организм осуществляет развитие по женскому типу. В результате появляется особь с кариотипом XY, но внешне более сходная с женщиной. Такие субъекты не способны иметь потомство, так как их половые железы (семенники) недоразвиты, а их выводные протоки часто формируются по женскому типу (недоразвитая матка, влагалище). Вторичные половые признаки также характерны для женского пола. Описанная картина известна у человека как тестикулярная феминизация, или синдром Морриса.Т,О достижение конечного результата при формировании у человека признаков мужского пола является следствием взаимодополняющего действия нескольких (по меньшей мере двух) генов, определяющих возможность синтеза мужского полового гормона и белка-рецептора. Др-м примером вз-я неаллельных генов служит формирование у человека групповой принадлежности крови по системе АВО. Выше этот признак рассматривался с точки зрения взаимодействия аллельных генов. Отвечающий за формирование данного признака ген I обеспечивает синтез антигенов А и В, фиксирующихся на поверхности эритроцитов. Однако для синтеза антигенов А и В необходимо образование неспецифического вещества-предшественника, которое детерминируется геном Н, находящимся в доминантном состоянии. У гомозигот по рецессивному аллелюhh даже при наличии в генотипе доминантных аллелей гена ^ или Iй антигены А и В не синтезируются и фенотипически эти люди имеют I группу крови. такая ситуация характерна для бомбейского феномена, описанного впервые в семье, где у женщины I группы крови родился ребенок с IV группой крови. Очевидно, ее фенотип был обусловлен непроявлениемимеющегося в генотипе аллеля IВ, что связано с гомозиготностью матери по рецессивному аллелюhh Вероятно с этих же позиций можно рассматривать и случаи альбинизма у человека, когда даже у африканских негров, которые обладают максимумом доминантных аллелей в системе полигенов, определяющих пигментацию кожи, появляются альбиносы с полным отсутствием пигмента в клетках кожи, радужке глаз, волосах.Альбинизм связывают с гомозиготностыо генотипа по рецессивному аллелю гена, не относящегося к полигенам пигментации. Возможно, его доминантный аллель определяет синтез продукта, принимающего участие в процессах, предшествующих синтезу пигмента. В таком случае здесь также имеет место взаимодополняющее действие соответствующих аллелей разных генов. В некоторых случаях при взаимодействии неаллельных генов для развития сложного признака необходимо обязательное присутствие одного из генов (А) в гомозиготном рецессивном состоянии (аа), тогда другой ген (В) обеспечивает формирование признака. Наличие в генотипе доминантного аллеля гена А каким-то образом препятствует проявлению гена В(b), и признак не формируется. Такое взаимодействие неаллельных генов принято называть эпистатическим Н:быть подавление у многих видов развития окраски покровов, определяемое одним из генов, при наличии в генотипе другого гена в доминантном состоянии. Так, у тыквы развитие окраски плодов определяется геном В. Доминантный его аллель детерминирует желтую, а рецессивный — зеленую окраску. Однако окраска не развивается вообще, если в генотипе имеется неаллельный ген А в доминантном состоянии. У кур породы леггорн белое оперение является результатом эпистатического воздействия доминантного аллеля одного гена на проявление неаллельного ему гена, отвечающего за пигментацию оперения. Отсутствие необходимых данных о роли первичных продуктов многих генов в формировании сложных признаков часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образования этих признаков. В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе. 16.Комбинативная изменчивость. Значение комбинативной изменчивости в обеспечении генотипического разнообразия людей. Адаптивный характер Комбинативной называют изменчивость, в основе которой лежит образование рекомбинаций, т. е. таких комбинаций генов, которых не было у родителей.Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами — пример комбинативной изменчивости. Взаимный обмен участками гомологичных хромосом, или кроссинговер. Он создает новые группы сцепления, т. е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей. Случайное сочетание гамет при оплодотворении. Благодаря этим процессам происх-ит «перестановка» генов и возн-ют новые комбинации.Все три источника комбинативной изменчивости действуют независимо и одновременноСами гены , структура и число хромосом при этом не изменяются , возникают только новые сочетания генов что приводит к появлению организмов с другими генотипами и фенотипами.У микроорганизмов , размножающихя бесполым путём , появились особые механизмы , приводящие к появлению комбинативной изменчивости - трансформация и трансдукция .Явление комбинативной изменчивости объясняет , почему в детях и внуках сочетаются признаки родственников по отцовской и материнской линии. Примерами комбинативной изменчивости являются все организмы , образующиеся в результате полового процесса. Значение комбинативной изменчивости:1Поставляет эволюционный материал ( до 80% ) для адаптациогенеза и формообразования в результате естественного отбора ( отмечалась Ч. Дарвиным в качестве важнейшего фактора видообразования ).2Материал для получения новых форм в селекции.ПРИМЕРЫ: 1.Форма рогов у косули, т.е у детей может быть разная форма рог.2. Рождение детей с 1 или 4ой группой крови гетерозиготных родителей, имеющие 2 и 3 группы крови.3 У цветка ночная красавица есть ген красного цвета лепестков А и ген белого цвета а. Организм Аа имеет розовый цвет лепестков. Модификационная изменчивостьНосит адекватный характерАдаптивные модификации расширяют возможности организма к выживанию и размножению в более широком диапазоне условий внешней среды. Возникающие в этих условиях наследственные изменения подхватываются естественным отбором и таким путем происходит более активное освоение новых экологических ниш и достигается более эффективная приспособляемость к ним.Н:У человека:1) увеличение уровня эритроцитов при подъеме в горы 2) увеличение пигментации кожи при интенсивном воздействии ультрафиолетовых лучей 3) развитие костно-мышечной системы в результате тренировок шрамы (пример морфоза) У насекомых и других животных:1) изменение окраски у колорадского жука вследствие длительного влияния на их куколки высоких или низких температур 2) смена окраски шерсти у некоторых млекопитающих при изменении погодных условий (например, у зайца) 3) различная окраска бабочек-нимфалид (например, Araschnialevana), развивавшихся при разной температуре У растений:1) различное строение подводных и надводных листьев у водяного лютика, стрелолиста и др. 2) развитие низкорослых форм из семян равнинных растений, выращенных в горах У бактерий: 1) работа генов лактозного оперона кишечной палочки (при отсутствии глюкозы и при присутствии лактозы они синтезируют ферменты для переработки этого углевода) 17.Классификация мутации. Спонтанные и индуцированные. Соматические и генеративные мутации. Мутации- внезапное, скачкообразное, наследуемое , стоикие изменения генетического материала, приводящее к изменению тех или иных признаков организма. Термин ввел генетик Д.Фриз1.По характеру изменения генетического аппарата: 1.Генные(точковые). 2.Геномные 3.Хромосомные. 2. Поспособу возникновения :1. Спонтанные-мутации, возникающие при нормальных условиях жизни. Спонтанный процесс зависит от внешних и внутренних факторов (биологические, химические, физические). Спонтанные мутации возникают у человека в соматических и генеративных тканях.2 Индуцированные мутации возникают при воздействии на человека мутагенами – факторами, вызывающими мутации. Мутагены же бывают трех видов: · Физические (радиация, электро – магнитное излучение, давление, температура и т.д.) · Химические (цитостатики, спирты, фенолы и т.д.) · Биологические (бактерии и вирусы)3.По отношению к зачатковому пути:1.соматические -мутации, возникающие в клетках тела и обусловливающие мозаичность организма, т. е. образование в нём отдельных участков тела, тканей или клеток с отличным от остальных набором хромосом или генов. Не передпо наследству н: 1 на кусте черной смородины м. появляться ветка с белыми ягодами. При вегет-ом разм-й нов.признак м. сохраняться и у потомков. Мичурин вывел новый сорт яблони –антоновку полуторафунтовую.2. люди у кот-ыхцвет 1го глаза отличается от цвета др-го.3.животные определенной масти, у которых на теле появ-ся пятна др-го цвета2.генеративные мутации- мутац в половых клетках, передается по насл-ву. Обнаруживается только в след-их поколениях. Примером генеративной доминантной мутации у человека может служить появление пузырчатости кожи стоп, катаракты глаза, брахифалангии (короткопалость с недостаточностью фаланг). Примером спонтанной рецессивной генеративной мутации у человека можно рассматривать гемофилию в отдельных семьях.4. По адаптивному занчени: Выделяют положительные, отрицательные и нейтральные мутации. Эта классификация связана с оценкой жизнеспособности образовавшегося мутанта.Вредные (патогенные) мутации приводят к гибели эмбриона (или плода) и к развитию наследственных и врождённых заболеваний. Нейтральные мутации обычно не влияют на жизнедеятельность организма (например, мутации, вызывающие веснушки, изменение цвета волос, радужной оболочки глаза). Благоприятные мутации повышают жизнеспособность организма или вида (например, тёмная окраска кожных покровов у жителей африканского континента).5. По локализации в клетке. Мутации делятся на ядерные и цитоплазматические. Плазматические мутации возникают в результате мутаций в плазмогенах, находящихся в митохондриях. Полагают, что именно они приводят к мужскому бесплодию. Причем такие мутации в основном наследуются по женской линии. 18.Хромосомные мутации связанные с измен-ем числа хромосом. Примеры наследс-ыхзаболевании. Нарушение расхождения бивалентов в анафазе I мейоза является причиной изменения количества хромосом в гаплоидном наборе гамет. Нерасхождение отдельного бивалента приводит к появлению одной гаметы, лишенной данной хромосомы, и другой, имеющей эту группу сцепления в двойном количестве Оплодотворение таких гамет нормальными половыми клетками приводит к появлению особей, в кариотипе которых изменено общее число хромосом за счет уменьшения (моносомия) или увеличения (трисомия) числа отдельных хромосом. Нарушения структуры генома, заключающиеся в изменении количества отдельных хромосом, называют анэуплоидией.Анеуплоидия — изменение (уменьшение — моносомия, увеличение — трисомия) числа хромосом в диплоидном наборе, т.е. не кратное гаплоидному (2n+1, 2n-1 и т.д.). Механизмы возникновения: нерасхождение хромосом (хромосомы в анафазе отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и «анафазное отставание» (в анафазе одна из передвигаемых хромосом отстаёт от всех других). Трисомия — наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау). Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0). В том случае, если в целом повреждается механизм распределения гомологичных хромосом между полюсами веретена (что наблюдается при его разрушении), клетка остается неразделившейся. Во второе деление мейоза она вступает не гаплоидной, а диплоидной. Из нее образуются диплоидные гаметы. Оплодотворение таких гамет приводит к образованию триплоидных организмов. Увеличение в кариотипе зиготы числа наборов хромосом называют полиплоидией(Зn, 4n, 5n и т.д.). 19Хромосомные мутации связанные с измен-ем структуры хромосом. Хромосомные мутации (аберрации) характеризуются изменением структуры отдельных хромосом. При них последовательность нуклеотидов в генах обычно не меняется, но изменение числа или положения генов при аберрациях может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма.Внутрихромосомные аберрации — аберрации в пределах одной хромосомы. К ним относятся делеции, инверсии и дупликации. Делеция— утрата одного из участков хромосомы (внутреннего или терминального), что может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития (например, делеция в регионе короткого плеча хромосомы 5, обозначаемая как 5р-, приводит к недоразвитию гортани, ВПР сердца, отставанию умственного развития). Этот симптомокомплекс обозначен как синдром кошачьего крика, поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье. Инверсия — встраивание фрагмента хромосомы на прежнее место после поворота на 180°. В результате нарушается порядок расположения генов. Дупликация — удвоение (или умножение) какого-либо участка хромосомы (например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития). Межхромосомные аберрации — обмен фрагментами между негомологичными хромосомами. Они получили название транслокаций. Различают три варианта транслокаций: реципрокные (обмен фрагментами двух хромосом), нереципрокные (перенос фрагмента одной хромосомы на другую), робертсоновские (соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч, в результате образуется одна метацентри-ческая хромосома вместо двух акроцентрических). Пример:Синдром «кошачьего крика» -делеция короткого плеча 5 хромосомы.Синдром Вольфа-Хиршхорна - делецией одного из сегментов короткого плеча 4й хромосомы. 20.Генные мутации связанные с изменением в структуре генов. Классификация моногенных заболевании. По характеру изменений в составе гена различают следующие типы мутаций: Делеции— утрата сегмента ДНК размером от одного нуклеотида до гена. Дупликации — удвоение или повторное дублирование сегмента ДНК от одного нуклеотида до целых генов. Инверсии — поворот на 180° сегмента ДНК размером от двух нуклеотидов до фрагмента, включающего несколько генов. Инсерции— вставка фрагментов ДНК размером от одного нуклеотида до целого гена. Трансверсии— замена пуринового основания на пиримидиновое или наоборот в одном из кодонов. Транзиции — замена одного пуринового основания на другое пуриновое или одного пиримидинового на другое в структуре кодона.Пример 1. Замена аминокислотного остатка в составе полипептида (миссенс–мутации). В состав молекулы гемоглобина человека входят две a–цепи (a–цепь закодирована в 16-ой хромосоме) и две b–цепи (b–цепь закодирована в 11-ой хромосоме). В состав b–цепи входит 146 аминокислотных остатков, при этом в нормальной b–цепи шестым аминокислотным остатком является глутаминовая кислота. С участием нормальной b–цепи образуется нормальный гемоглобин – HbA. В нетранскрибируемой нити участка ДНК, кодирующего b–цепь, глутаминовая кислота закодирована триплетом ГАА. Если же в результате мутации в ДНК произойдет замена триплета ГАА на триплет ГТА, то на месте глутаминовой кислоты в молекуле гемоглобина в соответствии с генетическим кодом появится валин. В итоге вместо гемоглобина HbA появится новый гемоглобин –HbS. Такая замена всего лишь одного нуклеотида и одной аминокислоты приводит к развитию тяжелого заболевания – серповидноклеточной анемии. На клеточном уровне эта болезнь проявляется в том, что эритроциты приобретают форму серпа и теряют способность к нормальному транспорту кислорода. ГомозиготыHbS/HbS умирают в раннем детстве. Зато гетерозиготыHbA/HbS характеризуются слабо измененными эритроцитами. При этом изменение формы эритроцитов значительно повышает устойчивость гетерозигот к малярии. Поэтому в тех регионах Земли, где свирепствует малярия (например, в Африке), отбор действовал в пользу гетерозигот. Таким образом, серповидноклеточная анемия – это пример относительности «полезности» и «вредности» мутаций. 2.Фенилкетонурия – нарушения превращения фенилаланина в тридин из-за редкого снижения активности фенилаланингидроксилазы 21. Понятие онтогенеза и филогенеза. Биологический закон как основа для понимания взаимоотношений онто и филогенеза эволюционных преобразований Онтогенез – это индивидуальное развитие организма, в ходе которого происходит преобразование его морфофизиологических, физиолого-биохимических, цитогенетических и этологических (у животных) признаков. Онтогенез включает две группы процессов: морфогенез и воспроизведение (репродукцию). В результате морфогенеза формируется зрелая особь. В эволюционном аспекте рассматриваются следующие явления онтогенеза: эмбриональные адаптации; филэмбриогенезы; эмбрионизация онтогенеза, автономизация онтогенеза. Термин «филогенез», или «филогения» используют для обозначения исторического развития живых организмов: как всего органического мира Земли, так и отдельных таксонов (от царств до видов). Термин «филогенез» ввел Э. Геккель в 1866 г. |