Шпоры по ветеринарной генетике. 1. Предмет и методы генетики
 Скачать 1.02 Mb. Скачать 1.02 Mb.
|

34. хромосомная теория наследственности.33. Генетическое док-во кроссинговера.36. Кроссинговер.37. Картирование хромосом.Сущность соматического кроссинговера заключается в том, что он осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей. Кроссинговер происходит между двумя несестринскими хроматидами гомологичных хромосом. У гетерозиготных особей наблюдаются отклонения в проявлении нормальных "признаков. Явление соматического кроссинговера было предсказано А. С. Серебровским в 1922 г. при анализе причин появления исключительных перьев у кур. В 1936 г. соматический кроссинговер обнаружил К. Штерн у дрозофилы. Он исследовал самок серых с нормальными щетинками, но гетерозиготных (АаВЬ) по рецессивным генам желтой окраски тела (о) и опаленных щетинок (Ь). На теле некоторых серых с нормальными щетинками мух наблюдались двойные пятна. Половина пятна желтая с нормальными щетинками и половина серая, но с опаленными щетинками. Появление двойных пятен К. Штерн объяснил митотическим кроссинговером, в результате которого образуется часть клеток, гомозиготных по желтой окраске тела (аа), и часть, гомозиготных по опаленным щетинкам (Jbb). Эти клетки становятся родоначальницами при образовании участков тела с желтой окраской и нормальными щетинками и с нормальной серой окраской и опаленными щетинками. В этом случае проявляется действие рецессивных генов, оказавшихся в гомозиготном состоянии. Таким образом, осуществление кроссинговера в соматических клетках ведет к появлению мозаиков. Кроссинговер иногда происходит и на стадии размножения при образовании половых клеток, когда гонии еще имеют диплоидное число хромосом. В этом случае процент кроссоверных гамет может быть очень высоким. Частота митотического кроссинговера ниже мейотического, однако его также можно использовать для генетического картирования. Соматический кроссинговер имеет место у животных, растений и человека. Факторы, влияющие на кроссинговер. На кроссинговер могут заметно влиять условия внешней среды и генотипические факторы. Обнаружены гены, выполняющие роль запирателей кроссинговера, и гены, повышающие его частоту. В третьей хромосоме дрозофилы выявлена мутация, которая прекращает процесс крос-синговера во всех парах хромосом. В качестве запирателей крос-синговера могут выступать некоторые перестройки хромосом. Чаще всего это бывает связано с инверсией (переворачиванием) того или иного участка в одной из гомологичных хромосом. На частоту кроссинговера могут влиять радиация, химические мутагены, концентрация солей, гормоны, лекарства. В большинстве случаев при воздействии этих факторов частота перекреста повышается. Нормальный перекрест хромосом может изменяться в зависимости от температуры, возраста, пола особи. Так, у тутового шелкопряда кроссинговер идет только у самцов и не бывает у самок. У дрозофилы кроссинговер наблюдается только у самвк, однако оказалось, что при рентгеновском облучении можно вызвать его и у самцов. У мыши кроссинговер бывает у обоих полов, но интенсивнее у самок; у голубей — у обоих полов, но чаще у самцов. В гетерохроматических, в частности прицентромерных, районах хромосом частота перекреста снижена, и истинное расстояние между генами на этих участках может быть изменено. Карты хромосом После того как была установлена связь генов с хромосомами и обнаружено, что частота кроссинговера всегда вполне определенная для каждой пары генов, расположенных в одной группе сцепления, встал вопрос о пространственном расположении генов в хромосомах. На основе анализа генетических исследований Т. Морган и его ученик А. Стертевант выдвинули гипотезу линейного расположения генов в хромосоме. Изучение взаимоотношений между тремя генами при неполном сцеплении показало, что частота (процент) перекреста между первым и вторым, вторым и третьим, первым и третьим генами равна сумме или разности между ними. Так, в одной группе сцепления расположены 3 гена — А, В, С. Оказалось, что процент перекреста между генами АС равен сумме процентов перекреста между генами АВ и ВС, частота перекреста между генами АВ оказалась равной АС — ВС, а между генами ВС = АС — АВ. Приведенные данные соответствуют геометрической закономерности в расстояниях между тремя точками на прямой. На этом основании был сделан вывод: гены расположены в хромосомах в линейной последовательности на определенных расстояниях друг от друга. На основании анализа частоты кроссинговера между генами к настоящему времени для многих видов животных и растений построены карты хромосом. Картой хромосом называется план расположения генов в хромосоме. Кестл провел опыт анализирующего тригибридного скрещива- ния кроликов с тройными рецессивами с целью выяснения сцепления между такими генами: сплошная окраска — С, гималайская окраска — с*1; белый жир — Y, желтый жир — у; черная окраска — В, коричневая окраска — Ъ. В результате анализирующего скрещивания было получено 908 кроликов восьми разных фенотипов соответственно количеству разных сортов гамет (табл. 4). Численное соотношение особей разных фенотипических классов указывало на отсутствие" независимого наследования по этим трем парам аллелей. Нужно было установить порядок расположения этих генов в хромосоме. Поскольку известно, что численность гамет родительских форм должна значительно превышать численность кроссоверных гамет, то можно прийти к выводу, что родительские комбинации генов были с*]КВ и СуЪ (276 + 275 = 551). Они составляли от общего числа 60,7 %. Далее при анализе исходим из того, что двойных перекрестов должно быть значительно меньше, чем одинарных. Меньше всего было комбинаций с**уВ и CYb(7 + 16 = = 23) — 2,5 %. Генотипы этих кроликов отличались отродитель-ских только тем, что Yи у поменялись местами. Так могло произойти только при двойном перекресте, и это является подтверждением того, что расположение генов было именно таким. Вычисляем частоту одиночных перекрестов. От одиночного перекреста да первом участке образовались гаметы CYBи chyb(рис. 15). Случаев одиночного перекреста на первом участке бтыло 101 (55 + 46), или 11,1 %. В результате одиночного перекреста на втором участке образовались гаметы cfiYbи Су В и получено особей 233 (125 + 108), или 25,7 %. Для того чтобы определить более правильно частоту одиночных перекрестов, мы должны к каждому из них прибавить величину двойного перекреста — 2,5 %, так как двойной перекрест проходил по обоим участкам хромосомы. Следовательно, частота кроссинговера на 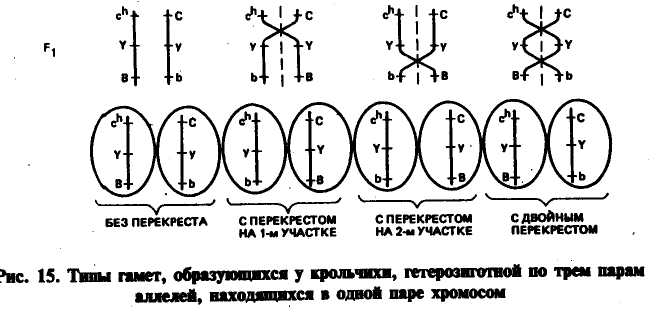 первом участке между генами с* и у составит 13,6 % (11,1 + 2,5). На втором участке между генами у и Ь — 28,2 % (25,7 + 2,5). Отсюда общая протяженность обоих участков, или процент перекреста между генами с* и Ъ составит 41,8 (13,6 + 28,2). Расстояние между генами с* и bможно определить и путем учета общего числа одиночных перекрестов (без включения двойных перекрестов). Оно составляет 36,8 %. Прибавив к этому удвоенный процент двойных перекрестов, т. е. 5,0 % (2,5 ■ 2), получим 41,8 %, что совпадает с результатами уже сделанного расчета по сумме перекрестов на каждом из участков. Теперь можно проверить, насколько фактическая величина двойного перекреста совпадает с теоретически ожидаемой. Теоретически ожидаемую величину рассчитывают путем перемножения процентов перекреста между генами на первом и втором участках, т. е. (13,6 : 100)(28,2 : 100)100 = 3,83 %. Фактически их было 2,5 %. Уменьшение числа ожидаемых двойных кроссоверов показывает, что кроссинговер на одном участке влияет на прохождение обмена на соседнем участке. Явление торможения кроссинговера на одном участке кроссинго-вером на другом получило Название интерференции. Чем меньше будет расстояние, разделяющее три гена, тем больше интерференция. Принимая во внимание линейное расположение генов в хромосоме, взяв за единицу расстояния частоту кроссинговера, Морган с сотрудниками составили первую карту расположения генов в одной из хромосом дрозофилы. Затем были составлены карты других ее хромосом. Оказалось, что установленное распределение генов в хромосоме является общебиологической закономерностью. К настоящему времени составлены карты хромосом для животных и растений многих видов. Если для какого-то вида установлена группа сцепления, которая содержит три и более гена, можно составить план их расположения в хромосоме. Так, в разобранном выше примере кроссинговер между генами с* и у обнаружен у 13,6 % кроликов, между генами у и Ь — у 28,2, а между генами с" и Ъ с учетом двойного перекреста — у 41,8 % животных. Ген Ъ не может быть расположен между генами с* и у, так как расстояние его от гена с* значительно больше, чем между генами с* и у (41,8 % против 13,6 %). Следовательно, три изученных гена расположены в хромосоме в таком порядке: Внизу цифрами указано расстояние между генами. Далее устанавливают сцепление хотя бы одного из этих генов с каким-то четвертым геном и снова проводят анализирующее скрещивание, выявляя частоту кроссинговера между вновь изучаемым геном и прежними хотя бы двумя уже изученными. На основании величины кроссинговера определяют его место в отношении к известным генам. При построении карт в хорошо изученных хромосомах указывают не расстояние между генами, а расстояние до каждого гена от нулевой точки начала хромосомы. Для лучшего усвоения материала нами везде доминантный аллель обозначается прописной буквой, рецессивный — строчной. В специальной литературе и при построении карт хромосом название (символ) гена может включать от 1 до 4 букв. Берется такое минимальное количество букв, чтобы сокращенное обозначение не совпало с любым другим обозначением генов данного вида организмов. При сокращении символа гена, как правило, ' берут первую букву названия признака и в случае необходимости добавляют буквы из середины или конца слова либо из других слов, входящих в название. Для обозначения генов рекомендуется использовать прилагательные или существительные латинского или английского языка. Например, гены курицы обозначают: Barring — В (оперение полосатое); Crest — Сг (черепно-мозговая грыжа); Prenatal — pn(гибель на поздней стадии инкубации). После построения генетических карт встал вопрос о том, отвечает ли расположение генов в хромосоме, построенное на основании частоты кроссинговера, истинному расположению. С этой целью генетические карты нужно было сравнить с цитологическими. В 30-х годах нашего столетия Пайнтер открыл в слюнных железах дрозофилы гигантские хромосомы, строение которых можно было изучать под микроскопом. Хромосомы эти имеют характерный для них поперечный рисунок в виде дисков разной толщины, которые окрашиваются кармином. Каждая хромосома по длине имеет специфические рисунки дисков, что позволяет отличать разные ее участки друг от друга. Появилась возможность сверить генетические карты с фактическим расположением генов в хромосомах. Материалом для проверки служили хромосомы, у которых вследствие мутации возникали различные хромосомные перестройки: или не хватало отдельных дисков, или они были перевернуты, или удвоены и т. д. Диски служили маркерами, с их помощью определяли характер хромосомных перестроек и место расположения генов, о существовании которых было известно на основании данных генетического анализа. При сопоставлении генетических карт хромосом с цитологическими было установлено, что каждый ген находится в определенном месте (локусе) хромосомы и что гены в хромосомах расположены в определенной линейной последовательности. В то же время было обнаружено, что физические расстояния между генами на генетической карте не вполне соответствуют установленным цитологически. Однако это не снижает ценности генетических карт хромосом для предсказания вероятности появления особей с новыми сочетаниями признаков. На основании анализа результатов многочисленных экспериментов с дрозофилой Т. Морган сформулировал хромосомную теорию наследственности, сущность которой заключается в следующем: 1) гены находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга; 2) гены, расположенные в одной хромосоме, относятся к одной группе сцепления. Число групп сцепления соответствует гаплоидному числу хромосом; 3) признаки, гены которых находятся в одной хромосоме, наследуются сцепленно; 4) в потомстве гетерозиготных родителей новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами; 5) на основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом. |