Шпоры по ветеринарной генетике. 1. Предмет и методы генетики
 Скачать 1.02 Mb. Скачать 1.02 Mb.
|

29. Летальные гены.Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот мэжет быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству. Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития. По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % —сублетальными (полулетальными) и менее 50 % — субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2—3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32—35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4—5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды. Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так: Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д. 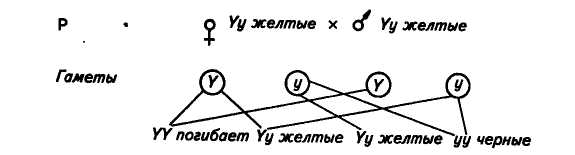 Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2—4-й день после рождения от уродства прямой кишки — отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина — гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли. (Более подробно об аномалиях при летальных генах будет сказано в соответствующей главе.) 35. Генетический анализ полного сцепления.30. Сцепленное наследование признаков32. Особенности наследования признаком при неполном и полном сцеленииСЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ ПРИЗНАКОВ Мы познакомились с дигибридным скрещиванием и уяснили, что независимое комбинирование признаков объясняется тем, что расщепление одной пары аллельных генов, определяющих соответствующие признаки, происходит независимо от другой пары. Однако это наблюдается только в том случае, когда гены разных пар находятся в разных парах хромосом и при образовании половых клеток гибрида в мейозе отцовские и материнские хромосомы независимо комбинируются. Но количество хромосом очень ограниченно по сравнению с количеством признаков, каждый из которых развивается под контролем определенного гена. Так, у дрозофилы известно около 7000 генов при четырех парах хромосом. Предполагается, что у человека не менее 50 тыс. генов при 23 парах хромосом, и т. д. Отсюда следует, что в каждой паре хромосом должны быть локализованы сотни аллелей. Естественно; что между генами, которые находятся в одной хромосоме, наблюдается сцепление и при образовании половых клеток они должны передаваться вместе. Сцепленное наследование открыли в 1906 г. английские генетики У. Бэтсон и Р. Пеннет при изучении наследования признаков у душистого горошка, но они не смогли вскрыть причины этого явления. Природу сцепленного наследования в 1910 г. выяснили ученые Т. Морган и его сотрудники К. Бриджес и А. Стертевант. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной для генетических опытов. В клетках тела дрозофилы находится 4 пары хромосом. Она отличается очень высокой плодовитостью — одна пара дает более ста потомков. У нее большая скорость развития — в течение 12—15 дней после оплодотворения из яйца развиваются личинка, куколка и взрослая особь, которая почти сразу же способна давать потомство. Можно исследовать в течение года более двадцати поколений. Мухи серого цвета, с красными глазами,. имеют маленькие размеры (около 3 мм), легко разводятся в биологических пробирках; для изучения их признаков можно пользоваться лупами. При просмотре сотен тысяч особей Морган обнаружил множество разных мутаций: встреча-лись мухи с черным и желтым телом, с белыми и другого цвета глазами, с измененной формой и положением крыльев и т. д. Иногда попадались особи, имеющие сразу несколько мутаций, например черное тело, зачаточные крылья, киноварные глаза. Изучая наследование разных пар признаков при дигибридном и полигибридном скрещиваниях, Морган и его сотрудники обнаружили большое число примеров сцепленного (совместного) их наследования. Все изученные признаки распределились на четыре группы сцепления в соответствии с числом и размерами хромосом у дрозофилы. На этом основании Морган сделал вывод о том, что гены, определяющие эти признаки, находятся в хромосомах. Гены, расположенные в одной хромосоме, представляют собой группу сцепления. Сцепление генов — это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом. Например, у дрозофилы 4 группы сцепления, у человека 23, у крупного рогатого скота 30, у свиней 19 и т. д. Мысль о расположении генов в хромосомах высказал Сеттон еще в 1902 г. Он обнаружил параллелизм в поведении хромосом в мейозе и наследовании признаков у одного из видов кузнечика. Дальнейшие исследования, проведенные Морганом, показали, что сцепление генов, расположенных в одной хромосоме, может быть полным или неполным. Наиболее четко разница в поведении сцепленных и независимо наследующихся генов выявляется при проведении анализирующего скрещивания. При независимом наследовании двух пар признаков у гибрида Fi (АаВЬ) с равной вероятностью образуется 4 сорта гамет: АВ, Ab, aB, ab. При скрещивании с полным рецессивом (aabb) количество сортов гамет у гибрида обусловливает число типов потомков и одинаковую вероятность их появления, так как гаметы рецессивной особи (ab) не могут изменить проявления доминантных и рецессивных генов гамет гибрида. В результате соотношение фенотипов потомства будет равно 1:1:1:1. Если же обе пары аллельных генов расположены в одной паре хромосом, то при образовании половых клеток гены этих аллелей не смогут свободно комбинироваться. В этом случае наблюдается сцепленное наследование. ПОЛНОЕ СЦЕПЛЕНИЕ Т. Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость — над зачаточными крыльями. Обозначим ген серой окраски тела В, аллельный ему ген черной окраски тела Ь; ген длиннокрылое™ V, аллельный ему ген зачаточных крыльев v. Обе пары этих генов нахо-дятся в одной и той же второй паре хромосом. По обеим парам Признаков родительские формы были гомозиготны: самка по рецессивному признаку черного тела (bb) и доминантному признаку длиннокрылое™ (VV), самец по доминантному признаку серой окраски. (ВВ) и рецессивному признаку зачаточных крыльев (w>. Гаметы родителей при редукционном делении получают у материнской формы хромосому с генами bи V, у отцовской"—с генами В и v. Все потомство первого поколения (Fi) имело серое тело и длинные крылья (рис. 13) и было гетерозиготно по обеим парам признаков (bV/Bv). Затем из Fi были отобраны самцы, которых скрестили с гомозиготными по обоим рецессивным генам самками, черными зачаточнокрылыми (bv/bv), т. е. было проведено анализирующее скрещивание, в результате которого при независимом комбинировании признаков должны были бы получить потомство четырех фенотипов в равных соотношениях: серых длиннокрылых, серых с зачаточными крыльями, черных длиннокрылых, черных с зачаточными крыльями, но были получены потомки только двух фенотипов, похожих на исходные родительские формы: черные длиннокрылые и серые короткокрылые. В этом случае наблюдается полное сцепление признаков. Это связано с тем, что у гетерозиготного самца в одной и той же хромосоме из гомологичной пары расположены и ген черной окраски, и ген длинных крыльев, в другой — ген серой окраски и ген зачаточнокрылости. При спермиогенезе в период мейоза гомологичные хромосомы расходятся в разные половые клетки. Образуется только два сорта гамет: один с хромосомой, которая несет гены Ъ и V, другой с хромосомой, в которой расположены гены В и v. При сочетании указанных гамет с гаметами особи с рецессивными признаками и образуется потомство только двух типов. При полном сцеплении гены, расположенные в одной хромосоме, всегда передаются вместе. Полное сцепление пока установлено только у са>щов дрозофилы и самок тутового шелкопряда. НЕПОЛНОЕ СЦЕПЛЕНИЕ В следующем опыте, так же как и в предыдущем, Морган скрещивал черных длиннокрылых самок с серыми зачаточно-крылыми самцами. В первом поколении получил все потомство серое длиннокрылое. Затем снова произвел анализирующее скрещивание, но из первого поколения отобрал не самца,, а самку и скрестил ее с черным с зачаточными крыльями самцом (рис. 14). В этом случае появилось потомство не двух типов, как при полном сцеплении, а четырех: серое о-зачаточными крыльями, черное длиннокрылое, серое длиннокрылое и черное с зачаточными крыльями, но не в равных соотношениях, как при независимом комбинировании признаков, а со значительным преобладанием фенотипов, сходных с родительскими формами. 41,5 % мух было серых с зачаточными крыльями, как у одного исходного родителя, и 41,5 % особей черных длиннокрылых, как у другого исходного родителя. Только 17 % потомков родилось с новым сочетанием признаков: 8,5 % черных с зачаточными крыльями и 8,5 % серых длиннокрылых. Таким образом, 83 % потомков имели сочетание признаков, как у исходных родительских форм, но появились особи и с новым сочетанием признаков. Следовательно, сцепление является неполным. / Встал вопрос: почему появились особи с новой комбинацией родительских признаков? Для объяснения этого явления Морган использовал и развил теорию хиазмотипии бельгийского цитолога Янсенса. В 1909 г. Янсенс наблюдал, что при спермиогенезе у саламандры в профазе мейоза гомологичные хромосомы конъ-югируют, а затем, при начале расхождения, образуют фигуры в хромосомы обмениваются участками. Если сцепленные гены лежат в одной хромосоме и у гетерозигот при образовании гамет происходит рекомбинация этих генов, значит, гомологичные хромосомы во время мейоза обменялись своими частями. Обмен гомологичных хромосом своими частями называется перекрестом или крос-синговером (английское слово crossingover означает образование перекреста). Особей с новыми сочетаниями признаков, образовавшимися в результате кроссинговера, называют кроссоверами. Вернемся к рисунку 14. У самки Fi, гетерозиготной по обеим парам признаков, в одной из гомологичных хромосом расположены гены Ъ и V, в другой — аллельные им гены В и v. В профазе редукционного деления, когда две гомологичные хромосомы соединились в один бивалент, каждая из хромосом удвоена и состоит из двух хроматид. Всего будет 4 хроматиды. Между двумя хроматидами гомологичных хромосом и происходит обмен их частями. В результате ген Ъ, расположенный в хроматиде одной гомологичной хромосомы, может соединиться с геном v, расположенным в хроматиде другой гомологичной хромосомы, и как результат одного события образуется вторая хроматида, где соединятся гены В и V. В дальнейшем хроматиды разойдутся и образуются кроссоверные гаметы с хромосомами с новым сочетанием генов (bvи BV). Две другие хроматиды из пары гомологичных хромосом не участвуют в перекресте и сохраняют в первоначальном сочетании материнские (bV) и отцовские (Bv) гены. Образование новых кроссоверных гамет обеспечило появление дрозофил -с новым сочетанием признаков: черных с зачаточными крыльями и серых длиннокрылых. Однако большая часть потомков будет сходна с исходными родителями (черные длиннокрылые и серые коротко-крылые). Морган приходит к выводу, что количество появления новых форм зависит от частоты перекреста, которая определяется по следующей формуле: Если, например, общее число потомков 900, а новых кроссоверных форм 180, то частота перекреста будет составлять 20 %. Морган установил, что частота перекреста между определенной парой генов — относительно постоянная величина, но различная для разных пар генов. На основании этого был сделан вывод о том, что по частоте перекреста можно судить о расстояниях между генами. За единицу измерения перекреста принята его величина, равная 1 %. Иногда ее называют морганидой. Величина перекреста зависит от расстояния между изучаемыми генами. Чем больше отдалены гены друг от друга, тем чаще происходит перекрест; чем ближе они расположены, тем вероятность перекреста меньше. Установлено, что количество кроссоверных особей к общему числу потомков никогда не превышает 50 %, так как при очень больших расстояниях между генами чаще происходит двойной кроссинговер и часть кроссоверных особей остается неучтенной. Их можно учесть при изучении не двух пар сцепленных признаков, а трех или четырех. В этом случае, учитывая двойные и тройные перекресты, можно точнее судить о расстояниях и частоте перекреста между генами. |