ответы 50-99. 50. Окислювальне фосфорилювання
 Скачать 370.05 Kb. Скачать 370.05 Kb.
|

|
церабразиди і гангліозиди Гексозою частіше є галактоза. Вуглеводна частина може бути не тиілки моносахаридом, а й олігосахаридом, який включає залишки галактозу, глюкози, ацетильованих глюкозамінів i галактозамінів, ряд інших. Таким чином, гліколіпіди є вуглеводними похідними церамідів. Вуглеводна частина гліколіпідів мембран бере участь у розпізнаванні клітинами молекул й вінших клітин. Зокрема, глікоцераміди є антигенами А i В мембран еритроцитів. Велика кількість цереброзидів відкрита у мембранах нервових клітин. Вони характеризуються специфічним набором жирних кислот: лігноцеринова, нервонова, церебронова. Частина моносахаридів у цереброзидах мозку сульфатована, тобто до одного із гідроксилів приєднаний залишок сірчаної кислоти. Ганглюзиди за будовою аналогічні до цереброзидів, але їх олігосахаридний ланцюг обов'язково включає один або декілька залишків N-ацетилнейрамінової кислоти. Гангліозиди знаходяться переважно у сірій речовині мозку, в плазматичній мембрані нервових i гліальних клітин. Гангліозидом є рецептор холерного токсину в кишечнику. 80.Перетравлення жирів у шлунково-кишковому тракті. Расщепление жира на глицерин и высшие жирные кислоты осуществляется под влиянием фермента липазы. Для воздействия липазы на жир необходимо его предварительное эмульгирование, достигаемое путем перемешивания в кишечнике пищевой кашицы с желчью. В ротовой полости жиры не подвергаются химическим изменениям. В желудке присутствует липаза, однако ее активность невелика из-за отсутствия условий, необходимых для эмульгирования жира. В желудке гидролизуются только эмульгированные жиры — жиры молока и яичного желтка. В основном переваривание жира происходит в кишечнике и в первую очередь в двенадцатиперстной кишке, куда по протокам попадают вместе с желчью соли желчных кислот, обладающие мощным эмульгирующим действием. Желчные кислоты образуют тончайшую пленку на жировых каплях, которая препятствует слиянию отдельных капелек жира в более крупные капли. Это приводит к резкому увеличению поверхности соприкосновения жира с ферментом липазой и, следовательно, скорости гидролитического распада жира. К желчным кислотам относятся холевая, дезоксихолевая и другие. По своему строению они близки к холестерину. В желчи эти кислоты образуют с глицином (гликоколлом) или таурином парные соединения — глико- или таурохолевую, глико- или тауродезоксихолевую и другие желчные кислоты, присутствующие в виде натриевых солей. В клетках кишечного эпителия из продуктов гидролиза пищевых жиров вновь ресинтезируются жиры, или липоиды, специфичные для данного вида животных. Синтезированные липиды транспортируются в жировые депо. При необходимости из жировых депо жиры могут переходить в кровь и использоваться тканями в качестве энергетического материала. МЕХАНИЗМ ОКИСЛЕНИЯ НЕЙТРАЛЬНОГО ЖИРА В ТКАНЯХ Поступивший в клетки нейтральный жир под действием тканевых липаз расщепляется на глицерин и высшие жирные кислоты. В дальнейшем жирные кислоты и глицерин окисляются в тканях на СО2 и H2O, при этом освобождающаяся энергия накапливается в макроэргических связях АТФ. ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ В ТКАНЯХ. В основе современных представлений о распаде жирных кислот в тканях лежит теория b-окисления, выдвинутая впервые Кноопом в 1904 г. Согласно этой теории, окисление жирных кислот происходит у углеродного атома, находящегося в b-положении по отношению к карбоксильной группе, с последующим разрывом углеродной цепочки жирной кислоты между a- и b-углеродными атомами. В дальнейшем эта теория была уточнена и дополнена. В настоящее время установлено, что окислению жирных кислот в тканях предшествует их активация при участии коэнзима А и АТФ. Этот процесс катализируется ферментом тиокиназой. Активированная жирная кислота (ацилкофермента А) подвергается дегидрированию, в результате чего возникает двойная связь между a- и b-атомами углерода. Этот процесс протекает с участием ацилдегидрогеназ, которые в качестве простетической группы содержат ФАД. Затем к ненасыщенной кислоте (a, b-не насыщенному производному ацил-КоА) присоединяется молекула воды и образуется b-гидроксикислота (b-гидроксиацил-КоА) . Далее снова происходит процесс дегидрирования с образованием b-кетокислоты (b-кетоацил-КоА). Этот процесс катализируется ацилдегидрогеназами, коферментом которых является НАД+.И на последнем этапе b-кетоацил-КоА, взаимодействуя со свободным КоА, расщепляется на ацетил-КоА и ацил-КоА. Последний укорочен по сравнению с первоначальным на два углерода. Образовавшийся на последнем этапе ацетил-КоА сгорает в лимоннокислом цикле до СО2 и H2O. Оставшийся ацил-КоА подвергается далее аналогичному превращению до полного окисления. Полное окисление насыщенной жирной кислоты приводит к высвобождению значительного количества энергии. Например, при полном окислении пальмитиновой кислоты образуется 131 молекула АТФ. 81.Роль жовчі в травленні жиру. Роль жовчі в травленні Після прийому їжі білки, вуглеводи, жири, вітаміни і мінеральні солі разом з кров'ю потрапляють в печінку. В процесі переробки клітинами печінки ці речовини набувають нову хімічну будову. Далі, через нижню порожнисту вену вони надходять до всіх тканин і органам і перетворюються в нові клітини тіла. Їх частина залишається в печінці, утворюючи своєрідне депо. Клітини печінки постійно виробляють жовч. Вироблювана жовч виділяється в просвіт капілярів, з них через жовчні ходи вона надходить в жовчні протоки, які зливаються в області воріт печінки, утворюючи печінковий протік. З нього секрет потрапляє в загальний жовчний протік або в жовчний міхур (через протоки міхура). Потрапивши в просвіт дванадцятипалої кишки, він стає учасником процесу травлення, бере участь у зміні шлункового травлення на кишкове. Печінка виробляє жовч безперервно. Прийом їжі посилює її відділення через 3-12 хвилин. Стимулюють вироблення жовчі м'ясо, молоко, хліб, жовтки яєць. Властивості жовчі, що виробляється печінкою Жовч інактивує пепсин, нейтралізує кисле вміст шлунка і створює сприятливі умови для активної роботи ферментів підшлункової залози. Вона стимулює секрецію шлункового слизу, підшлункової залози, покращує моторну і секреторну діяльність тонкої кишки. Наявність в жовчі травних ферментів дозволяє приймати участь в процесі кишкового травлення, вона попереджає поява гнильних процесів. «Якість» жовчі визначається її основними компонентами. До них відносяться жовчні кислоти, холестерин, жовчні пігменти. Жовчні кислоти - це специфічні продукти обміну речовин в печінці, холестерин і жовчні пігменти мають внепеченочной походження. У клітинах печінки з холестерину утворюються первинні жовчні кислоти: холевая і хенодезоксихолева. Жовчні кислоти, що надійшли в кишечник, беруть участь у перетравлюванні і всмоктуванні жирів. Жовчні пігменти є продуктами метаболізму гемоглобіну, вони надають секрету характерну забарвлення. Жовч впливає на всмоктування в тонкій кишці жиророзчинних вітамінів (Д, Е, К), солей кальцію, холестерину, нерозчинних у воді жирних кислот. Вона стимулює моторну діяльність тонкої кишки, (в тому числі кишкових ворсинок), в результаті цього швидкість всмоктування речовин в кишці посилюється, приймає участь в пристеночном травленні - створює сприятливі умови для закріплення ферментів на поверхні кишечника. 82.Всмоктування ліпідів. Більшість ліпідів всмоктується в нижній частині дванадцятипалої і у верхній частині тонкої кишки, інші – в інших ділянках тонкої кишки. Продукти розщеплення ліпідів всмоктуються епітелієм ворсинок. Всмоктуюча поверхня епітеліальної клітини збільшена за рахунок мікроворсинок. Епітеліальна клітина в середньому містить до 3000 мікроворсинок. Кожна мікроворсинка має один субмікроскопічний каналець. Ліпідні речовини і продукти їх розщеплення проникають у порожнину клітини покривного епітелію двома методами: через субмікроскопічні канальці мікроворсинки і через інтерстеціальні щілини. В першому випадку процес відбувається за допомогою дифузії, осмосу і активного транспортування, в другому – завдяки піноцитозу, тобто захопленню клітинною поверхнею найдрібніших харчових частинок. У людини і тварин 10% жиру всмоктується у вигляді тригліцеридів, 10% – у ди- і моногліцеридів, 80% – у вигляді продуктів кінцевого гідролізу. Продукти перетворення ліпідів складаються з дрібних частинок жиру, ди- і моногліцеридів, вищих жирних кислот, гліцерину, гліцерофосфатів, азотистих основ, холестерину, вищих спиртів, фосфорної кислоти та ін. Вони розподіляються в двох фазах: ліпідній і міцелярній. У ліпідній фазі основними компонентами є найдрібніші частинки три- і дигліцеридів, у міцелярній – вищі жирні кислоти, моногліцериди та інші продукти перетворення ліпідів. Продукти перетворення ліпідів всмоктуються неоднаково. Просто всмоктуються гліцерин і гліцерофосфати, інозин і сфінгозин. Фосфорна кислота всмоктується у вигляді натрієвих і калієвих солей. Азотисті основи всмоктуються за участю нуклеотидів типу цитидиндифосфата, утворюючи комплекс, наприклад цитидин-дифосфатхолін: Вищі жирні кислоти не розчиняються у воді, але за наявності жовчних кислот утворюють розчинні комплекси – холеїнові кислоти. На одну молекулу вищої жирної кислоти в розчинних комплексах доводиться в середньому 2 – 4 молекули жовчних кислот. Співвідношення між ними в комплексах може бути наступним: 4:1, 7:2, 8:3, 9:3 і т.д. У розчинному комплексі гідрофобний радикал вищої жирної кислоти оточений з усіх боків молекулами жовчних кислот. Гідрофільна частина молекул жовчних кислот розміщується зовні розчинного комплексу і просто взаємодіє з водою. В итогі цього розчинний комплекс просто проникає в порожнину епітеліальної клітини. Найбільший ступінь всмоктування мають олеїнова і масляна кислоти (98 – 90%), менший – пальмітинова і стеаринова (88 – 60%). Стерини теж не розчиняються у воді. Їх всмоктування відбувається після взаємодії з жовчними кислотами і утворення холеїнових кислот. Багато стеринів, особливо рослинного походження, всмоктуються повільно, гальмуючи засвоєння організмом холестерину. При всмоктуванні окремих ліпідів спостерігається синергізм. Так, ступінь всмоктування жирів і холестерину зростає при збільшенні в кишках екзогенного і ендогенного лецитину. Останній бере участь в утворенні стійких емульсій і хіломікронів. Припускають, що лецитин здійснює перенесення гліцеридів через слизову оболонку кишки в кровоносне русло. У товстій кишці немає ферментів які б проявляли гідролітичну дію на ліпіди. Ліпідні речовини, які не зазнають змін в тонкій кишці, в цій ділянці харчового каналу піддаються гнильному розкладанню під впливом ферментів мікрофлори. Слиз товстої кишки містить деяку кількість фосфатидів. Частина з их резорбується. Холестерин, що не всмоктався, відновлюється до копростерина. 83.Окислення гліцерину. Глицерин увлекается преимущественно печенью. Здесь под действием глицеролкиназы он превращается в глицерофосфат, который окисляется до диоксиацетонфосфату глицеролфосфатдегидрогеназою. Диоксиацетонфосфат – промежуточный продукт гидролиза и глюконеогенеза и поэтому может или окисляться в реакциях гликолиза и далее по общему пути катаболизма до СО2 и Н2О, давая при этом энергию, или вступать в реакции глюконеогенеза, превращаясь в глюкозу или гликоген. Окисление глицерина в анаэробных условиях приводит к выделению двух молекул АТФ (как и в гликолизе), но учитывая, что одна молекула АТФ была использована для активации глицерина, энергетический баланс равный одной молекуле АТФ. При полном окислении глицерина в аэробных условиях до СО2 и Н2О энергетический баланс составляет 22 молекулы АТФ. Из них 9 АТФ образуются в дыхательной цепи с 3-х молекул НАДН2. Одна молекула НАДН2 – при окислении глицерофосфата, вторая – с глицеральдегидтрифосфату, а третья молекула НАДН2 образуется при преобразовании пировиноградной кислоты в ацетил КоА. Окисления последнего до СО2 и Н2О сопровождается выделением 12 АТФ. 84.Окислення вищих жирних кислот. Енергетичне значення процесу окислення жирних кислот. Жирова тканина, що складається з адипозоцитів, виконує специфічну роль у ліпідному обміні. Близько 65% маси жирової тканини припадає на частку відкладених у ній триацилгліцеролів (ТАГ) - вони являють собою форму запасання енергії та виконують в обміні жирів таку ж функцію, як глікоген печінки в обміні вуглеводів. Відкладені жири в жировій тканині служить джерелом ендогенної води і енергетичним резервом для організму людини. ТАГ використовується в організмі після попереднього розщеплення (ліполізу), в ході якого звільняються гліцерин і вільні жирні кислоти. У клітинах жирової тканини за участю ліпаз відбувається розпад ТАГ. Ліпаза знаходиться в неактивній формі, вона активується гормонами (адреналіном, норадреналіном, глюкагоном, тироксином, глюкокортикоїдами, СТГ, АКТГ) у відповідь на стрес, голодування, охолодження продуктами реакції є моноацілгліцерінов і ВЖК. ВЖК за допомогою альбумінів переносяться кров'ю до клітин тканин, органів, де відбувається їх окислення. Окислення вищих жирних кислот. Джерела ВЖК: - Ліпіди жирової тканини - Ліпопротеїни -Тріацілгліцеріни -Фосфоліпіди клітинних біомембран Окислення ВЖК відбуваються в мітохондріях клітин, і називається бетта окисленням. Доставка їх до тканин і органів відбувається за участю альбуміну, а транспорт з цитоплазми в мітохондрії за участю карнітину. Процес бета-окислення ВЖК складається з наступних етапів: -Активація ВЖК на зовнішній поверхні мембрани мітохондрій за участю АТФ, конзіма А та іонів магнію з утворенням активної форми ВЖК (ацил - КоА). -Транспорт жирних кислот всередину мітохондрій можливий при приєднанні активної форми жирної кислоти до карантину, що знаходиться на зовнішній поверхні внутрішньої мембрани мітохондрій. Утворюється ацил-карнітин, що володіє здатністю проходити через мембрану. На внутрішній поверхні комплекс розпадається і карнітин повертається на зовнішню поверхню мембрани. -Внутрімітохондріальное окислення жирних кислот складається з послідовних ферментативних реакцій. В результаті одного завершеного циклу окислення відбувається відщеплення від жірой кислоти однієї молекули ацетил-КоА, тобто вкорочення жирнокислотного ланцюга на два вуглецевих атома.Пріетом в результаті двох дегідрогеназну реакцій відновлюється ФАД до ФАДН 2 і НАД + до НАДН 2. 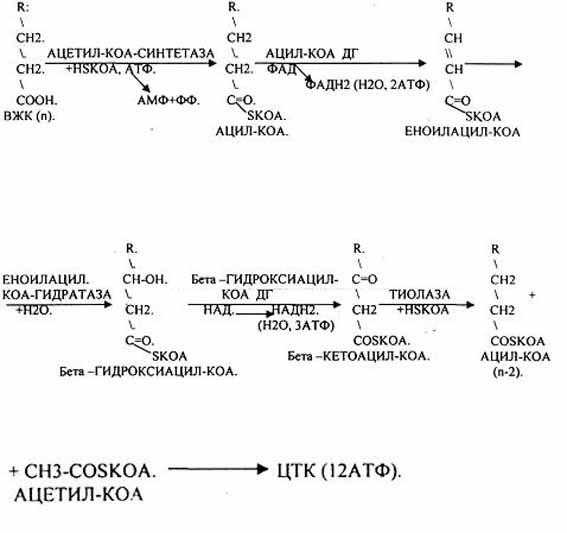 рис. Окислення вищих жирних кислот Т.ч. завершуючи 1 цикл бігу - окислення ВЖК, в результаті якого ВЖК вкоротити на 2 вуглецевих ланки. При бета-окисненні виділилося 5АТФ і 12АТФ виділилося при окисленні ацетил-КоА в ЦТК і зв'язаних з ним ферментів дихального ланцюга. Окислення ВЖК буде відбуватися циклічно однаково, але тільки до останньої стадії - стадії перетворення масляної кислоти (БУТІРІЛ-КОА), яка має свої особливості, які необхідно враховувати при підрахунку сумарного енергетичного ефекту окислення ВЖК, коли в результаті одного циклу утворюється 2 молекули ацетил-КоА , одна з них проходила бета-окислення з виділенням 5АТФ, а інша ні. рис. Остання стадія окислення вищих жирних кислот ^ ОКИСНЕННЯ ВЖК, що має непарний кількість вуглецевих ланкою в ланцюзі Такі ВЖК надходять в організм людини в складі їжі з м'ясом жуйних тварин, рослин, морських організмів. Окислення таких ВЖК відбувається також як і ВЖК, які мають парну кількість вуглецевих ланок у ланцюзі, але тільки до останньої стадії - стадії перетворення пропіонов-КОА. яка має свої особливості. 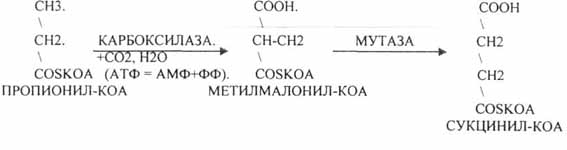 Т.ч. утворюється сукцініл-КоА, який в подальшому окислюється в мітохондріях за участю ферментів ЦТК Кребса і зв'язаних з ним ферментів дихального ланцюга. 85.Роль різних органів та тканин в обміні ліпідів. 86.Розщеплення та синтез фосфатидів Фосфолипиды стимулируют использование жиров в организме. При недостатке фосфатидов замедляются процессы биохимического превращения жиров в печени, и содержание их в этом органе может достигать 50%, вместо 5% в норме. При гидролитическом распаде фосфолипидов образуются глицерин, жирные кислоты, фосфорная кислота и азотистые основания. Первые два продукта могут окисляться до СО2 и воды или могут принять участие в синтезе жиров. Один из представителей азотистых оснований холин является продуктом распада лецитинов и принимает участие в синтезе ряда важных для организма соединений (метионин, креатин и др.). Ацилирование холина уксусной кислотой в организме приводит к образованию ацетилхолина, который имеет большое значение в передаче нервных возбуждений В молекуле лецитина, который является наиболее распространенным фосфатидом, фосфорная кислота соединена, например, с азотсодержащим спиртом — холином. Приведенная ниже формула строения лецитина и осо бенно схема его гидролитического распада дает исчерпывающее представление в его строении Как показывает опыт, жировое перерождение печени может быть предотвращено введением с пищей достаточного количества лецитина. Впоследствии выяснилось, что способностью предотвращать ожирение печени и даже удалять уже отложенный жир из печени обладают и другие вещества, в частности входящий в состав лецитина холин, а также аминокислота метионин. Причина отложения жира в печени при отсутствии холина в настоящее время до некоторой степени выяснена. Как указывалось, в печени постоянно происходит не только интенсивный распад фосфатидов, но и одновременный синтез их из нейтральных жиров. Для этого синтеза, помимо высших жирных кислот и неорганических фосфатов, необходимо наличие азотистого основания холина. Но при недостаточном образовании или недостаточном поступлении в печень уже готового холина синтез липоидов из жиров становится либо невозможным, либо резко задерживается, и нейтральный жир отлагается в печени. Метионин обладает способностью отдавать свою подвижную метильную группу, необходимую для синтеза холина (стр, 347). Этим и объясняется тот факт, что белок казеин, в состав которого входит большое количество метионина, также обладает липотропным действием, т. е. способствует удалению из печени избытка жира. Ацетальфосфатиды (плазмалогены) при гидролизе распадаются на глицерин, фосфорную кислоту, этаноламин (оксиэтиламин) или холин и альдегид высшей жирной кислоты (пальмитиновой или стеариновой). Ацеталь-фосфатидам приписывают следующее строение На приводимой схеме, на примере лецитина, можпо проследить ферментативный распад фосфатидов до их составных частей Обмен фосфатидов тесно связан с обменом жиров, однако, в отличие от жиров, биосинтез и распад фосфатидов в растениях изучены в значительно меньшей степени. Важная роль фосфатидов в протоплазме клеток, в обмене веществ и прежде всего в регуляции проницаемости протоплазмы для различных веществ заставляет в последнее время предпринять детальное изучение их биосинтеза и распада. В реакциях распада фосфатидов может принимать участие еще один фермент—фосфолипаза В, которая катализирует расщепление фосфатидов до фосфатидных кислот и свободных азотистых оснований. Реакция идет по следующей схеме [c.328] Фосфатиды. Фосфатиды, так же как и жиры, являются глицеридами, т. е. сложными эфирами глицерина и жирных кислот. От настоящих жиров они отличаются тем, что содержат фосфорную кислоту и связанное с ней азотистое соединение. Азотистые соединения в фосфатидах обычно представлены холином и кол-амином. При гидролизе фосфатиды распадаются на глицерин, фосфорную и жирные кислоты и на холин (или коламин). [c.324] Дальнейший обмен продуктов распада фосфатидов—высших жирных кислот и глицерина, был освещен ранее. Поэтому рассмотрим здесь лишь последующие превращения холина. [c.407] Вначале в зерне при повышении температуры интенсивно идут различные ферментативные процессы крахмал превращается в сахар, начинают расщепляться белки, и образуется некоторое количество аминокислот, а также могут гидролизоваться жиры и жироподобные вещества (например, фосфатиды). При повышении температуры до 70° ферментативные процессы прекращаются, но зато усиленно происходят реакции взаимодействия между продуктами распада указанных сложных веществ — моносахаридами, аминокислотами и органическими кислотами. В результате этих реакций и образуются вкусовые и красящие вещества, о которых [c.165] Липопротеиды. При гидролизе липопротеиды распадаются на белок и растворимые в эфире жиры, лецитины и другие фосфатиды. [c.503] Фосфатиды и слизи гидрофильны и поэтому снижают водостойкость покрытий Свободные жирные кислоты и продукты их распада снижают скорость высыхания покрытий, ухудшают их механические свойства Красящие вещества придают маслам интенсивную окраску, что затрудняет их использование для получения покрытий светлых оттенков Антиоксиданты замедляют окислительную полимеризацию — основной процесс при пленкообразовании масел Вследствие этого растительные масла, предназначенные для производства лакокрасочных материалов, подвергают очистке от вредных примесей [c.192] Опыты с радиоактивным фосфором (Р ) показали, что и обмен фосфатидов, их распад и синтез в нервной ткани также происходит относительно медленно, главным образом вследствие ограниченной скорости проникания неорганических фосфатов в головной мозг. Интенсивность обмена значительно снижается при инсулиновой коме, рассеянном склерозе и некоторых других патологических состояниях. [c.433] Липопротеиды. Это наименее исследованная группа белков. При гидролизе распадаются на белок и растворимые в эфире лецитины и другие фосфатиды, т. е. соединения, содержащие фосфорную кислоту, связанную с гидроксильными группами многоатомных спиртов (см. стр. 659). [c.712] К катализаторным ядам относятся каротиноиды, разрушающиеся в процессе гидрогенизации госсипол и его производные, фосфатиды, жирные кислоты, мыла и соединения серы. Госсипол и его производные необратимо отравляют катализатор, так как не гидрируются в обычных условиях и не распадаются на катализаторе. Свободные жирные кислоты и их мыла реагируют с металлом катализатора, образуя на нем прочно удерживаемый поверхностный слой никелевых (или медных) мыл, блокирующий активные центры катализатора. [c.185] Липопротеиды. Это наименее изученная группа белковых, тел. При гидролизе распадаются на белок и лецитины (фосфатиды). Липопротеиды содержатся в хлорофильных зернах, а также в протоплазме. [c.345] Распад фосфатидов, отщепление от них азотистых соединений, фосфорной кислоты или жирных кислот катализируется ферментами, которые получили название лецитиназ или фосфолипаз. Известно несколько типов таких ферментов. [c.311] Фосфатиды наиболее интенсивно распадаются в растениях при прорастании семян, однако механизм этих процессов выяснен недостаточно детально. По аналогии с животными тканями можно предположить, что на первом этапе от фосфатида (лецитина или кефалина) отщепляется одна жирная кислота, в результате чего образуется лизолецитин или лизокефалин. Эта реакция катализируется ферментом фосфолипазой А. При распаде лецитина реакция идет следующим путем [c.327] А. И. Смирнов установил, что в условиях фосфатного голодания в тканях этиолированных проростков гороха усиливается гидролиз полисахаридов и фосфатидов и активируется экзосмос продуктов распада через обедненную фосфатами протоплазму. При этом наблюдалось также замедление миграции азотистых соединений из семядолей в ткани проростков и подавление в последних процессов синтеза белков. На более поздних этапах фосфатного голодания происходил распад ранее образовавшихся в проростках белков. [c.416] Вопрос о том, как синтезируются ненасыщенные л<ирные кислоты, встречающиеся в составе жиров и фосфатидов, остается еще невыясненным. Также еще мало известно о пути их распада. Некоторым исключение г является олеиновая кислота, путь превращения которой изучен более полно. Опыты с введением в организм животных олеиновой кислоты, меченной тяжелым изотопом водорода (дейтерием), показали, что она превращается в стеариновую кислоту (т. е. гидрируется), которая затем подвергается Р-окислению. [c.316] Потребление фосфатидов при эмбриогенезе происходит у человека и у млекопитающих животных. Плодом по мере развития потребляются фосфатиды, доставляемые ему с кровью матери. Содержание фосфатидов в крови пупочных артерий оказывается в течение всего периода эмбриогенеза выше, чем в крови пупочных вен. Более детальные исследования показали, что фосфатиды, доставляемые к плоду с кровью матери, подвергаются там распаду, а из продуктов распада синтезируются фосфатиды тканей плода. [c.322] Образовавщиеся в результате распада фосфатидов свободные жирные кислоты, фосфатидные кислоты, глицеролфосфат и азотистые основания в зависимости от направленности процессов обмена веществ подвергаются в организмах тем или иным превращениям. [c.328] Холин, возникающий как при распаде фосфатидов, так и путем синтеза из серина и метионина, в некоторой своей части подвергается ацетили-рованию с образованием ацетилхолина, играющего важную роль в передаче нервных импульсов (стр. 568). [c.325] Обмен фосфатидов. Пути распада фосфатидов. Современные представления о путях распада фосфатидов в организме основаны главным образом на тщательном изучении превращений, которые свойственны фосфати-дам вне организма при воздействии на них теми или иными ферментами. Поэтому когда говорят о путях распада фосфатидов, то имеют в виду скорее возможные, чем действительные, пути их деструкции. Непосредственно в биологических объектах эти пути исследованы еще недостаточно. Однако известно, что время полужизни фосфатидилглицерина и дифосфатидилглицерина у бактерий составляет 1 и 2 ч соответственно, а период полужизни фос-фоинозитидов и сфингомиелинов в мозге крысы—12,5 и 40 суток соответет-венно. [c.405] Несмотря на то что фосфатиды в некоторой степени предотвращают прогоркание жиров, кремы с лецитинами должны быть консервированы, так как в условиях, благоприятствующих жизнедеятельности микроорганизмов, лецитин распадается с выделением холина НО СНг СН2Ы(СНз)зОН, который при дальнейшем распаде выделяет триметиламин Ы(СНз)з. [c.39] В поджелудочном соке находятся ферменты лецитиназа В, глицерофосфатаза> и холинфосфатаза (соответственно коламинфосфатаза). В связи с этим в кишечнике фосфатиды, подвергаясь гидролизу (лецитиназа В отщепляет одновременно обе жирные кислоты), распадаются на глицерин, жирные кислоты, фосфорную кислоту и холин (или коламин). [c.284] Опыты с радиоактивным фосфором (Р ) показали, что и обмен фосфатидов, их распад и синтез в нервной ткани также происходит очень медленно. Из фосфатидов мозга инозитфосфатиды (особенно инозитдифосфатид) наиболее активно вовлекаются в процессы обмена. [c.409] Гликолипиды состоят из остатков какого-нибудь сахара, глицерина и жирных кислот. Галактолипиды, имеющие в своем составе галактозу, являются основными липидами хлоропластов. На их долю приходится около /з от всего количества липидов этих органоидов. По всей вероятности, они так же, как фосфатиды и сульфолипи-ды, играют роль структурного материала ламелл хлоропластов. Некоторые считают, что помимо структурной роли, они могут выполнять функцию запасного вещества хлоропластов, так как содержат много сахаров и в определенных условиях, например в темноте, могут распадаться с освобождением сахаров. [c.72] Кроме главной составной части — смеси сложных эфиров, природные жиры всегда содержат примеси других веществ — остатки белков, углеводов и солей (из клеточного сока растений), стерины и стериды, воски, фосфатиды, витамины, ферменты и продукты их распада Растительные жиры могут еще содержать небольшие количества каротина, хлорофилла, ксантофилла, а также эфирных масел. Многие из перечисленных примесей мешают дальнейшей переработке жиров, способствуют их порче при хранении, и поэтому их необходимо предварительно удалять, что достигается очисткой, или так называемой рафинацией. [c.192] Как уже упоминалось, в процессе обмена веществ у растений и животных образуются преимущественно Р = 0-метаболиты II, VI и VII, из которых сульфоксид (VI) и сульфон (VII) значительно более устойчивы в водных системах растений и животных, чем не окисленный Р = О-метилдеметон (II) (табл. 8 и 9 на стр. 53, 54 демонстрируют большую гидролитическую устойчивость соединения VI по сравнению с метаболитом II гидролитическая устойчивость метаболитов VII и VI приблизительно одинакова [668]). Поэтому соединения VI и VII остаются некоторое время в растениях и в основном выполняют функцию носителей системного инсектицидного и акари-цидного действия. По мере того как они распадаются до диметилфосфорной кислоты (VIII), а затем до фосфорной кислоты и соответствующих спиртов, системное действие ослабевает. Продолжительность системного действия зависит от внешних воздействий и вида растения [894]. У картофеля, например, оно затухает через 12—13 дней после обработки препаратом [124]. Образующаяся при полном разложении метилдеметона фосфорная кислота принимает участие в построении растительных фосфатидов [669]. [c.107] Фосфатиды, поступающие с гшщей (в составе печени, яичного желтка, мозга и т. д.), в кишечнике подвергаются гидролитическому распаду под влиянием ферментов фосфолипаз. При этом разрываются эфирные связи между глицерином, жирными кислотами, фосфорной кислотой и холином (или коламином). Имеется несколько фосфолипаз, обозначаемых А-, В-, С- и О-фосфолипа-зы в зависимости от точки приложения их действия на молекулу фосфатида [c.408] Соединения этого типа широко распространены в живых организмах. К ним относятся глицеринфосфорная кислота, входящая в состав фосфатидов (стр. 88), триозо- и гексозо-фосфорные кислоты (стр. 280), являющиеся промежуточными продуктами распада углеводов, серинфосфорная кислота, содержащаяся в фосфопротеидах (стр. 41) и в некоторых фосфатидах (стр. 88), и другие. При гидролизе фосфорных эфиров освобождается небольшое ко.личество энергии (2500—3000 калорий на моль расщепленного эфира). [c.217] Липиды представляют собою большую группу органических веществ, различных по химической структуре и обладающих одним общим для них свойством — нерастворимостью в воде и растворимостью в различных органических растворителях. Вполне попят1ю, что путь превращения различных по своей структуре липидов, их распад и синтез ие люгут быть одинаковыми. Различной должна быть и роль их в организме. Ниже мы остановимся на обмене жиров, фосфатидов и стеридов. О роли липовита-минов и стеринов уже сообщалось (стр. 118 и 50). [c.303] До сих пор излагались данные об окислительном распаде насыщенных жирных кислот. Естественно возникает вопрос, как окисляются ненасыщенные жирные кислоты. На первый взгляд может показаться, что этот вопрос разрешается просто. Ведь известно, что при р-окислении, уже на первом его этапе, при дегидрировании, образуется из насыщенной кислоты ненасыщенная, поэтому можно предположить, что ненасыщенные жирные кислоты являются промежуточными продуктами окисления насыщенных жирных кислот. Этому, однако, противоречат следующие обстоятельства. Во-первых, при Р Окислении возникает ненасыщенная связь между атомами углерода в положенги а- и Р- по отношению к карбоксилу. Между тем как в жирах, так и в фосфатидах встречаются жирные кислоты, имеющие не одну, а несколько ненасыщенных связей. Даже в случае одной ненасыщенной связи, например, з олеиновой кислоте, она расположена далеко от карбоксила — между 9-м и 10-м атомами углерода. Во-вторых, некоторые жирные кислоты с несколькими ненасыщенными связями (линолевая, линоленовая и арахи-доновая) н( могут образоваться в организме животных из других веществ и должны доставляться с нищей. Эти жирные кислоты содержатся в растительных жирах и поэтому эти жиры биологически более ценны, чем жиры животного происхождения. [c.316] В превращениях фосфатидов в организме, в их распаде и синтезе участвует ряд ферментов. Исходя из того, что фосфатиды отличаются друг от друга по составу своих компонентов, следует полагать, что в превращениях их участвуют различные специфические для них ферменты. Наиболее изучены ферменты, активирующие распад холинфосфатидов (лецитинов) и эта-ноламинофосфатидов (кефалинов). [c.322] Печень — один из важнейщих органов, играющих активную роль в процессах обмена веществ. Продукты переваривания пищи, а также вещества, возникающие в результате жизнедеятельности микрофлоры кишечника, всасываются в кровь и по системе воротной вены (vena porta hepati a) доставляются в печень. В печени питательные вещества подвергаются различным превращениям. В печени происходит синтез сложных органических веществ белков, гликогена, фосфатидов и других соединений. В ней же сложные органические молекулы (гликоген, жирные кислоты) подвергаются распаду с образованием сравнительно несложных по своей структуре веществ, которые поступают в кровь и кровью доставляются к различным тканям и органам, где и используются. [c.482] |