Ксенобиология ответы. 6. Реакции биотрансформации неорганических ксенобиотиков
 Скачать 1.37 Mb. Скачать 1.37 Mb.
|

|
Примеры: 1) В Англии создана автоматизированная система биологического контроля качества воды с помощью рыб. В проточный аквариум помещается форель. При подаче загрязненноц воды под влиянием поллютантов она ослабевает и сносится в задний отсек, где стоит фотоэлемент, возникает сигнал опасности. 2) Наиболее удачными для использования в качестве индикаторов являются споровые и кокковые микроорганизмы. Это позволяет получить информацию о содержании ряда КБ с учетом процессов их трансформации в природных обьектах. В качестве тест реакций – ростовые параметры, ферментативные реакции. 14. Простая диффузия ксенобиотиков через биологические мембраны. Ионные каналы. При простой диффузии ксенобиотики проходят через мембрану в результате случайного молекулярного движения, и величина потока линейно зависит от концентрации и коэффициента проницаемости мембраны для данного вещества. Биологические мембраны представляют собой мозаичную структуру. «Сердцевина» мембраны представляет собой в основном гидрофобную область, поэтому неполярные вещества сравнительно легко проникают в клетки. Гидрофобные группы способствуют увеличению проникающей способности, полярные – ее уменьшению. Скорость диффузии обусловлена стерическими факторами ( если разветвлена углеводородная цепь- уменьшается скорость). В качестве количественного параметра проникновения любого ве-щества в клетку вводится коэффициент проницаемости. В этом случае скорость переноса определяют с помощью закона Фика:  где D – коэффициент диффузии; А – площадь поверхности; Cн, Cв – концентрация снаружи и внутри соответственно; Кр – коэффициент распределения, представляющий отношение концентрации растворенного вещества в мембране к концентрации вне мембраны; Dx – расстояние, преодолеваемое соединением при прохождении через мембрану. Так как молекуле, возможно, приходится преодолевать путь Dx, не обязательно равный толщине мембраны, то этот путь трудно измерить непосредственно. Коэффициент распределения обычно определяют, используя в качестве липидной фазы оливковое масло, а не истинные липиды, что вводит некоторую неопределенность в величину К. Зависи-мость D от x также неизвестна ни для какой мембраны. Поэтому Р = DKр/Dx (коэффициент проницаемости) является единственной измерямой величиной, характеризующей мембрану и переносимое растворенное вещество, которую легко вычислить, используя выражение: dS/dt = PA(Cн – Cв), Р – длина в единицу времени (т. е. скорость), см/с. Проникающая способность некоторых веществ через мембрану зависит от коэффициента распределения. От-ношение концентраций какого-либо вещества, состоящего из простых молекул, в двух находящихся в равновесии фазах имеет постоянное зна-чение и называется константой распределения Кр: Кр = С2/С1, где С1 – водная фаза, С2 – неводная фаза (масло, липид) – справедливо в том случае если нет взаимодействий между растворенным веществом и растворителем Пассивный перенос – это движение вещества по градиенту электро-химического потенциала без затраты энергии. Движущая сила- электрохимический потенциал и он обладает свободной энергией вещества. Химический потенциал определяется условным уровнем энергии. Если через мембрану переносятся незаряженные частицы, то Если система в состоянии равновесия и нет градиента электрохимического потенциала, то вещество проходит из-за теплового движения беспорядочного. В таком случае два противоположно направленных потока сбалансированы RTlnCjн + ZjF 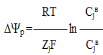 – Потенциал Нернста. Проницаемость мембран для некоторых малых молекул оказывается более высокой, чем это можно было бы ожидать на основании данных об их растворимости в липидах. Хорошо проникают через мембраны такие гидрофильные вещества, как глицерин, мочевина и др., радиус молекул которых не превышает 0,3 нм. Вероятно эти молекулы проходят через участки мембраны= поры( каналы) либо заряженные частицы проникают через ионные каналы. Канал – это белковая макромолекула, субъединицы которой обра-зуют в мембране пору через бислой липидов. В поре имеется узкий селективный фильтр вблизи наружной поверхности мембраны и воротное устройство вблизи ее внутренней поверхности. Пространство между селективным фильтром и воротами получило название туннеля. Сенсор напряжения, расположенный в липидном слое, управляет открытием ворот под влиянием внутримембранного поля. Расширенные части канала у наружной и внутренней поверхности образуют устья (или вестибюли). 15.Влияние физиологических и генетических факторов на биотрансформацию ксенобиотиков. К генетическим факторам относятся видовые различия и различия внутри одного вида. Видовые различия и различия внутри одного вида. Различия процессов биотрансформации между видами могут быть количественными (идинтичные реакции протекают с неодинаковой скоростью) и качественными (различные метаболические реакции). Качественные различия особенностей реакции можно продемонстрировать на следующем примере: у собак не происходит ацетилирования ароматических аминов, у кошек нет N-ацетилтрансферазы, у морских свинок не образуются меркаптанконьюгаты. Пример количественного различия.Так, в процессе деметилирования антипирена препарат печени хомяка почти в 200 раз активнее препарата печени краба. Видовые различия в действии ксенобиотиков (лекарственных веществ) связан, например с особенностями реакций коньюгации у разных животных. Классическим примером служит коньюгация фенилуксусной кислоты. У человека и шимпанзе фенилуксусная кислота коньюгирует с глутамином, у большинства других млекопитающих – с глицином и глюкуроновыми кислотами, а у кур – с орнитином. Кролики поедают листья белладонны безнаказанно, так как в их сыворотке содержится неспецифическая эстераза,способная гидролизовать алкалоиды группы тропина, тогда как у других млекопитающих этот фермент отсутствует. Различия между линиями внутри одного вида как у животных, так и человека. Хорошо известен пример генного полиморфизма при ацетилировании ксенобиотиков. Причина этого явления в различной активности N-ацетилтрансферазы – фермента, катализирующего реакции коньюгации ариламинов с ацетил-КоА. У некоторых людей ацетилирование протекает медленно, их называют «медленными ацетиляторами», а у других – «быстрых ацетиляторов» – в несколько раз быстрее. Было обнаружено, что полиморфизм ацетилирования наблюдается для ксенобиотиков, молекула которых трансформируется путем присоединения по атому азота ацетильной группы с участием N-ацетилтрансферазы: RH3 + CH3-CO КоA → RNH-COCH3 + HS-КоА Данный феномен определяется генетическими факторами. Активность N-ацетилтрансфкразы контролируется двумя аллелями одного локуса. К физиологическим факторам, которые влияют на метаболизм, относятся возраст, пол, состояние питания, заболевания и т. д. Возраст. Процесс развития характеризуется резким увеличением активности ферментов, в том числе и отвечающих за метаболизм ксено-биотиков. Это является фактором адаптации новорожденных к новым условиям существования. У новорожденных отсутствуют микросомальные ферменты, в том числе и цитохром Р-450. Их появление происходит в течение первых дней после рождения, и достигает максимума пример-но через 30 дней (4 недели) у крыс, через 8 недель у человека. Таким образом, эмбрионы и новорожденные особенно чувствительны к токсикантам. Способность новорожденных синтезировать коньюгаты также заметно снижена. Пол. У взрослых самцов чужеродные соединения метаболизируется быстрее, чем у взрослых самок. Это обусловлено действием половых гормонов на синтез ферментов микросомного окисления, т. к. эффект проявляется только при достижении половой зрелости и исчезает при ка-страции животных. Ци-тохром Р-450 состоит из набора изоэнзимов, а содержание некоторых из них зависит от пола. Питание и диета. Активность энзимов метаболизма ЧС отчетливо зависит от питания животного. У мышей голодание приводит к уменьшению скорости гидроксилирования одних ксенобиотиков и увеличению других. У крыс, находящихся на диете с дефицитом белка, наблюдается уменьшение активности ферментов моноксигеназных систем. Гормоны. Введение крысам тироксина вызывает уменьшение актив-ности ферментов монооксигеназной системы. Напротив, стероидные гормоны стимулируют активность микросомных ферментов, в первую очередь благодаря индукции их синтеза. Беременность. В конце беременности заметно уменьшается глюку-ронидная конъюгация ксенобиотиков, вероятно, из-за наличия в тканях протестерона – ингибитора глюкуронилтрансферазной активности в печени и других тканях. 16.Влияние факторов окружающей среды (стресс, температура, химические соединения и др.) на биотрансформацию ксенобиотиков. К факторам окружающей среды можно отнести неблагоприятные условия, вызывающие стресс, облучение ионизирующей радиацией, тем-пература и т. д., наличие других ксенобиотиков, а также большое влияние на процессы (скорость) метаболизации оказывает природа (структура) самих ксенобиотиков. Стресс. Неблагоприятные внешние условия приводят к стрессовым состояниям и увеличению микросомального окисления, зависящего от НАДФН, т. е. интенсивность метаболического превращения повышается. Ионизирующая радиация вызывает типичную стрессовую ответную реакцию. Ионизирующая радиация подавляет образование НАДН и НАДФН, и поэтому возможно нарушение микросомального окисления в печени. Ионизирующая радиация приводит к угнетению гидроксилирования стероидов. Температура. Скорость течения различных биологических процессов изменяется в зависимости от изменения температуры. Температурный коэффициент Q10 показывает насколько меняется скорость (V) того или иного процесса при изменении температуры на 10 оС. Q10 = Vt/V(t-10), Q10 для физико-химических процессов находится в интервале 1,1–1,5, химических реакций – 2–3, а для отдельных биохимических про-цессов и выше. Интенсивность биотрансформации ксенобиотиков возрастает при предварительном охлаждении экспериментальных животных, при этом в крови у них возрастает уровень гормона щитовидной железы (индуктор метаболизма ксенобиотиков), а в печени - содержание цитохрома Р-450. Как правило, процессы метаболического превращения в значительной степени зависят от температуры окружающей среды для ксенобио-тиков, влияющих на температуру тела. Так, токсичность ФОС (понижают температуру тела) для крыс снижается при содержании животных в условиях компенсируемой гипертермии (до 30 оС). Чужеродные соединения. Активирование метаболизма чужеродных соединений введением других ксенобиотиков, таких как медикаменты, пестициды и полициклические углеводороды, хорошо известно и широко изучается в связи с отношением этого феномена к лекарственному си-нергизму и толерантности, а также индукции ферментов и канцерогенезу. Ксенобиотики оказывают стимулирующий эффект путем увеличе-ния количества микросомальных ферментов, в том числе цитохрома Р-450 и НАДФН-цитохром редуктазы. Для соединений, стимулирующих микросомальные ферменты, характерна высокая растворимость в липидах и медленная скорость метаболизма. Полярные соединения не стимулируют микросомальные ферменты. Однако может наблюдаться и снижение активности цитохрома Р-450 в живых организмах. Чужеродное вещество либо связывается с гидрофобными сайтами (конкурентное ингибирование), либо взаимодействует с железом геминовой группы( неконкурентное ингибирование). Соединения, разрушающие мембрану ЭР, в которую встроены ферменты биотрансформации также их ингибируют. Ингибирующий и стимулирующий эффекты ксенобиотиков часто приводят к изменению токсичности и фармакологической активности. Так же есть зависимость скорости метаболических превращений от природы вещества. Скорости метаболизма микросомами печени крысы ряда многоядерных ароматических углеводородов: пирен, хризен, бензо–[а]–антрацен и др. Основная реакция в их метаболических превращениях – это реакция гидроксилирования. В результате гидроксилирования, сопровождаемого введением молекулы воды, образуются дигидролы. Даже для ряда близких по структуре многоядерных углеводородов скорость метаболизма изменяется более чем в двадцать раз. Пирен и хризен метаболизируются с меньшей скоростью по сравнению с другими исследованными веществами. 17. Основные пути поступления и выведения гидрофильных и гидрофобных ксенобиотиков живыми организмами. Ксенобиотик, который тем или иным образом попадает в организм, чаще всего подвергается метаболическому превращению с последующим его выведением. Основные пути поступления токсических ксенобиотиков в организм человека – это органы дыхания и кожа, а также пищеварительный тракт. Через слизистую оболочку дыхательных путей и пищ. тракта поступают ксенобиотики, находящиеся в газо- и парообразном состоянии. Через кожу проникают преимущественно липофильные вещества, находящиеся в жидком или газообразном состоянии при непосредственном контакте. Через всасывание из пищеварительного тракта. Участок всасывания (слизистая оболочка желудка, тонкой или толстой кишки) определяется физико-химическими свойствами ксенобиотика, его способностью к ионизации, взаимодействию с мембранами эпителиацитов. Экскреция ксенобиотикови их метаболитов через почки, легкие, кожу, кишечник, слюнные, потовые, слезные, сальные железы, а также молочные железы при лактации. 1)Почечная экскреция ксенобиотиков – основной путь удаления гидрофильных ксенов. В основе почечной экскреции лежат следующие биологические процессы: клубочковая фильтрация, канальцевая секреция, канальцевая реабсорбция. В клубочковой фильтрации подвергаются вода, глюкоза, аминокислоты, белки, ксенобиотики-неэлектролиты. Канальцевая секреция – активный процесс, осуществляемый с помощью ферментных систем мембранного транспорта (органические кислоты или органические основания). Канальцевая реабсорбция – процесс обратного всасывания метаболитов и ксенобиотиков, неионизированные формы веществ подвергаются реабсорбции и экскреции путем пассивной диффузии. 2) Экскреция ксенобиотиков легкими. удаляются в основном летучие и газообразные вещества, например промышленные газы, а также продукты печеночной биотрансформации многих токсических веществ.3) Экскреция ксенобиотиков печенью. Выделение через систему желчных ходов или после обратного всасывания в синусоиды – через почки. Однако для многих полярных ксенов (метаболитов, конъюгатов) выделение в желчь из гепатоцитов осуществляется путем активного транспорта против градиента электрохимического потенциала. У растений нет спец. путей поступления и выведения ксенобиотиков (через корни и листья, а строение корня опр. пути и механизмы поступления). Поступление в корни происходит в две фазы: 1) быстро протекающая диффузия в апопласт и 2) продолжительное поступление. Поступает широкий спектр гидрофильных и липофильных органических молекул ксенобиотиков (спирты, фенолы, амины). Ксены поступают в растение через листья: через устьица или кутикулу. Через кутикулу соединения диффундируют медленнее, чем через устьица. Однако из водных растворов поступление через устьица неэффективно, т.к. у входа в устьичную щель выпуклый мениск препятствует поступлению экзогенного соединения. Кутикула проницаема для гидрофильных и липофильных молекул. В распределении ксенобиотиков в тканях и органах растений большую роль играют сосуды ксилемы и флоэмы. Ксены, переносимые по сосудам ксилемы поступая в корни растений создают фототоксические конц-и в наземных органах растений. редко отмечается акк-я флоэмнобильных ксенов в корнях при поступлении их через листья (из-за выдел-я ксенов в ризосферу). Выделение через листья очень редко. 18. Описание основных процессов поведения ксенобиотиков в экосистемах. Роль адсорбции и перемещения. Для ксенобиотиков, попавших в экосистемы и входящие в них организмы, можно выделить следующие этапы. 1. Реакции превращения ксенобиотиков( распад, ОВР, гидролитические и реакции конъюгации. 2. Адсорбция ксенобиотиков на частицах биологического и абиотического происхождения. 3. Переход ксенобиотиков из одной среды в другую. Реализация этих этапов в реальных биогеоценозах происходит при теснейшем взаимодействии между различными факторами. Для понимания характера воздействия ксенобиотиков на экосистемы чрезвычайно важное значение имеет скорость превращения ксенобиотиков. Высокая скорость превращения обычно приводит к исчезновению вещества и, следовательно, к исчезновению проблемы, связанной с загрязнением окружающей среды; при медленном разрушении вещество сохраняется длительное время, что может вызвать его концентрирование. Адсорбция ксенобиотиков на частицах. Доступность ксенобиотика для ферментов,и его «деградабельность» снижается в результате сорбции молекул на частицах. Процессы сорбции–десорбции ксенобиотиков нередко определяют их устойчивость и взаимосвязаны с переносом в биосфере на большие расстояния. Пример: многие пестициды в почве гидролизуются с образованием соединений, которые адсорбируются на частицах почвы, связываются с гумусом и сохраняются длительное время, т. е. возрастает устойчивость этих токсикантов. В адсорбированном состоянии они не разрушаются. По мере разрушения гумуса грибами (что является ферментативным процессом) ранее связанные молекулы пестицида освобождаются и могут проявить свое токсическое действие на организмы данной экосистемы. Переходы веществ из одной среды в другую.Изменения физико-химических свойств веществ в результате модификации структуры их молекул, сорбция ксенобиотиков на частицах и др. оказываются существенными при переходе веществ (и продуктов их превращений) из одного блока биогеоценоза в другой. Таковы переходы ксенобиотиков из воды в воздух и обратно, из организмов в воду и обратно, из почвы в воду и т. д. Подобные переходы могут иметь решающее значение для крупномасштабного перемещения ксенобиотиков в атмосфере и порождает экологические проблемы. Например, летучесть ряда пестицидов (особенно хлорорганических), их переход в результате испарения из почвы или воды в воздух обусловливает их дальнейший перенос на большие расстояния. Однако не менее серьезные проблемы возникают и вследствие затрудненности перехода подобного типа. Например, переход ртути из почвы в воду происходит очень медленно (период полувыведения из почвы в поверхностные воды составляет 850 лет). Попавшая в водоемы ртуть далеко не безвредна: она метилируется и накапливается в гидробионтах. Поэтому возникает огромное запаздывание в миграции ртути по биосфере, которое затрудняет борьбу с ртутным загрязнением. В самом деле, даже если удалось бы сегодня полностью прекратить антропогенное загрязнение биосферы (в частности, почвы) ртутью, то все равно еще сотни лет будет продолжаться ее выход (вымывание) из почвы в водоемы, а следовательно, их загрязнение и последующее накопление в гидробионтах. |