Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

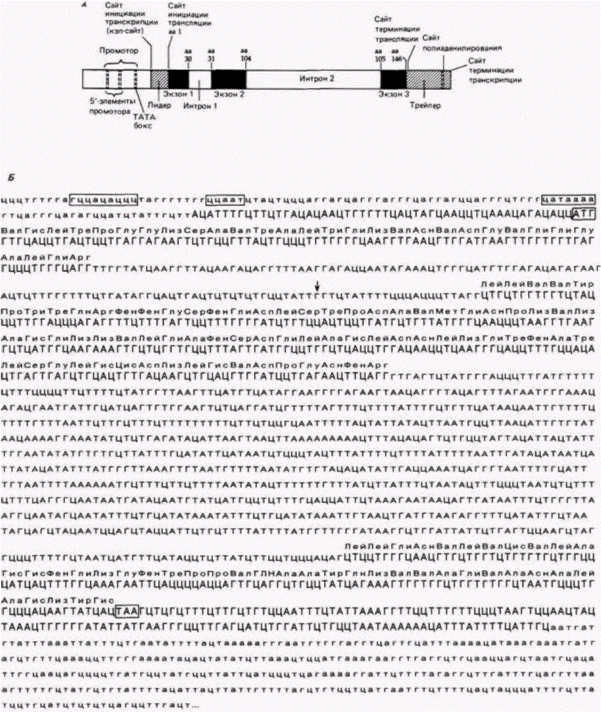
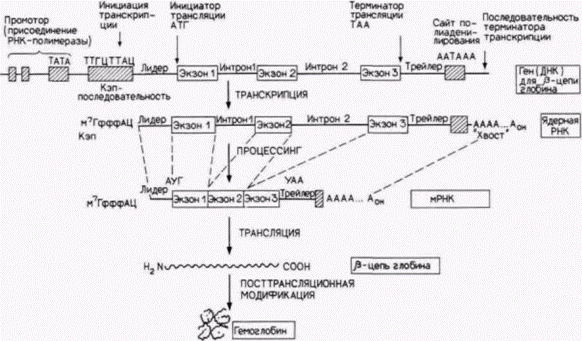
Глава 12. Регуляция экспрессии генов на уровне транскрипции: механизмы дифференциальной транскрипции геновВ чем бы ни заключалась конкретная работа генов, их функционирование можно отнести с уверенностью к категории процессов развития и. следовательно, к области эмбриологии. В настоящее время эта центральная проблема фундаментальной биологии атакуется со многих сторон как физиологами, так и генетиками; но в сущности она является эмбриологической проблемой. К. УОДДИНГТОН (1956) Мы вошли в клетку, нашу колыбель, и начали составлять опись обретенного нами богатство. АЛЬБЕР КЛОД (1974) ЭУКАРИОТИЧЕСКИЕ ГЕНЫ, КОДИРУЮЩИЕ БЕЛКИЭкзоны и интроныОдно из самых удивительных открытий семидесятых годов, десятилетия, отмеченного выдающимися открытиями, – выяснение структуры эукариотического гена. Генетики, изучающие бактерий, пришли к выводу, что белковый продукт колинеарен кодирующему его гену, т.е. 5’-конец РНК определяет аминоконцевую аминокислоту белка, и каждый кодон транслируется в аминокислоту до тех пор. пока не будет достигнут карбоксильный конец белка. В 1977 г. было обнаружено, что в случае некоторых мРНК эукариот дело обстоит иначе. Большинство расшифрованных с тех пор эукариотических генов оказались разорванными. Иными словами, 5'- и 3'-концы информационной РНК не происходят из одного непрерывного участка хромосомы. Между участками ДНК, кодирующими белок (экзонами), лежат промежуточные последовательности (интроны), которые не имеют никакого отношения к аминокислотной последовательности белка1. На рис. 12.1 показана структура гена ß-цепи гемоглобина человека. Этот ген состоит из следующих элементов: 1. Область промотора, ответственная за присоединение РНК-полимеразы и последующую инициацию транскрипции. Эта область содержит, как будет показано позже, три различных элемента и располагается с 95-й по 26-ю пару оснований перед сайтом инициации транскрипции (т.е. от –95-й до —26-й). 2. Последовательность АЦАТТТГ, на которой инициируется транскрипция. Ее часто называют кэп-последовательностью. потому что она кодирует 5'-конец РНК, который получит вскоре после транскрипции группу из модифицированных нуклеотидов – кэп (о чем будет говориться ниже). 3. Кодон АТГ, инициирующий трансляцию. Этот кодон лежит через 50 пар оснований после точки инициации транскрипции. Промежуточная область из 50 пар оснований между точками инициации транскрипции и трансляции называется лидерной последовательностью. 4. Экзон, содержащий 90 пар оснований, которые кодируют аминокислоты 1–30 в ß-цепи гемоглобина человека. 1 Известны, однако, интересные исключения (например, гены гистонов), когда таких промежуточных последовательностей в генах не содержится. Любая гипотеза, касающаяся функции этих последовательностей, должна учитывать такие исключения. Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ______________________________ 139
5. Интрон из 130 пар оснований, который не содержит последовательностей, кодирующих гемоглобин. 6. Экзон, содержащий 222 пары оснований, которые кодируют аминокислоты 31–104. 7. Большой интрон – 850 пар оснований, не вносящий вклада в белковую структуру гемоглобина 8. Экзон, содержащий 126 пар оснований, которые кодируют аминокислоты 105 – 146. 9. Кодон терминации трансляции – ΤАА. 10. Транскрибируемая 3’-концевая область, которая не транслируется в белок. Эта область включает последовательность ААТААА, которая необходима для присоединения к РНК-транскрипту «хвоста» приблизительно из 200-300 адениловых остатков. Этот поли(А)-конец вставляется в РНК примерно через 20 оснований после ААУААА-мотива. Однако прежде, чем транскрипция завершается, она продолжается после ААТААА-сайта на расстояние около 1000 нуклеотидов. В случае если в ААТААА-сайте возникла какая-либо мутация, поли(А)-хвост не добавляется и транскрипция не останавливается (Logan et al.. 1987). В пределах транскрибируемой 3'-концевой области расположена последовательность энхансера (примерно 600– 900 пар оснований от ААТААА-сайта), которая необходима для временной и пространственной организации экспрессии ß-глобинового гена в предшественниках зрелых эритроцитов (Trudel, Constantini, 1987). Изначальный ядерный РНК-транскрипт для такого гена содержит кэп-последовательность, лидерную последовательность, экзоны, интроны и 3'-нетранслируемый участок (рис. 12.2). Кроме того, оба конца транскрипта претерпевают модификацию. Концевая группа, названная кэп-группой (от англ. cap, что значит «шапочка»), состоящая из метилированного гуанозина, помещается на 5'-конце РНК. Таким образом, в отличие от предшественника мРНК, все нуклеотиды которого связаны в направлении от 5' к 3', кэп-структура присоединена в направлении от 5' к 5'. Это означает, что в ядерной РНК отсутствует свободная 5'-фосфатная группа (рис. 12.3). Молекулы зрелой информационной РНК содержат аналогичные кэп-группы, хотя остается неясным, действительно ли кэп-группа в мРНК является той самой, что была изначально получена в ядре. Известно также, что концевая 5'-кэп-группа необходима для присоединения мРНК к рибосоме (Shatkin, 1976). 3'-Конец обычно модифицируется в ядре путем присоединения хвоста примерно из 200 остатков аденилата. Эти остатки адениловой кислоты соединяются друг с другом ферментативно и добавляются к транскрипту. Они не кодируются последова- |