Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.140_______________ ГЛАВА 12______________________________________________________________________
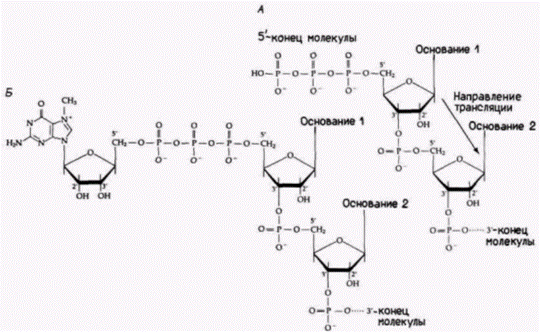
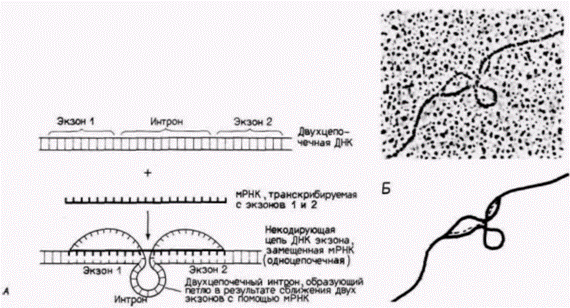
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ_________________________ 141 тельностью гена. Обе модификации, 5'- и 3'-концов, могут защищать РНК от экзонуклеаз (Sheiness, Darnell, 1973; Gedamu, Dixon, 1978), стабилизируя таким образом мРНК и ее предшественник. Эти поли(А)хвосты прогрессивно укорачиваются по мере старения мРНК. Новообразованная глобиновая мРНК в клетках мыши и кролика имеет около 150 адениловых остатков, тогда как старые РНК – 100, 60 или 40 аденилатов (Merkel et al., 1975; Nokin et al., 1976). Показано, что глобиновая мРНК, лишенная поли(А), быстро разрушается после инъекции ее в ооциты Xenopus. Глобиновая мРНК с поли(А)хвостом существует в этих условиях более 20 ч (Marbaix et al., 1975). Наличие интронов между кодирующими областями было выявлено с помощью электронного микроскопа (рис. 12.4). Информационная РНК ß-цепи глобина связывается со специфическими участками ядерного гена, выпячивая интроны. Имеются разного рода данные, что эти интроны необходимы для того, чтобы последовательности мРНК покинули ядро и попали в цитоплазму, где они могут быть транслированы в белки. Во-первых, можно сконструировать искусственные вирусы так, что в них будет встроена часть глобинового гена. Затем этим вирусам дают возможность инфицировать клетки и транскрибировать РНК в ядрах Если вирус содержит интрон (из глобинового гена или из вирусного гена), то в цитоплазме обнаруживаются стабильные последовательности глобиновой мРНК. Однако если интрон вируса или глобинового гена отсутствует, то последовательности глобиновой мРНК в цитоплазме не накапливаются (Hamer, Leder, 1979). Во-вторых, как уже было отмечено ранее, некоторые индивидуумы страдают от болезни, называемой β+-талассемией; это заболевание характеризуется недостаточной продукцией ß-цепей глобина. У больных имеются гены для ß-цепей глобина, и транскрипция предшественников мРНК на этих генах проходит, по-видимому, нормально. Более того, мРНК этих больных правильно транслируется в аминокислотную последовательность ß-цепи глобина человека. Дело заключается в количестве ß-глобиновой мРНК. Следовательно, заболевание проявляется на уровне процессинга РНК. Это было показано с помощью пульсового мечения («pulsechase») (Maquat et al., 1980). Развивающиеся эритроциты, выделенные из костного мозга больных, в течение 12 мин инкубировали с радиоактивными нуклеотидами («pulse»). Затем транскрипцию останавливали актиномицином D. Через различные интервалы времени («chase») из клеток выделяли РНК, которую подвергали электрофорезу для разделения молекул РНК по размерам, и затем гибридизовали эти РНК с ß-глобиновой кДНК. У нормальных индивидуумов вскоре после мечения появляется РНК длиной 1900 оснований. Несколько позже кДНК связывается с молекулами РНК, содержащими 1550, 1150, 960 и 880 оснований. Еще несколькими минутами позже 85% всей РНК представляет собой мРНК. В клетках от больных ß+-талассемией наблюдается переизбыток промежуточных продуктов. Очень незначительная часть этих РНК имеет к 30 минутам размеры зрелой мРНК. Образовавшиеся промежуточные продукты разрушаются, вероятно, в ядре, и лишь немногие выходят в цитоплазму (Maquat el al., 1980; Kantor et al., 1980). Таким образом, β+-талассемия обусловлена дефектом в процессинге предшественника ß-глобиновой мРНК. В результате секвенирования ß-глобиновых генов нескольких больных было обнаружено, что они содержат мутации в первом интроне. Мутация в одном из этих генов показана на рис. 12.1 (Spritz et al., 1981). Итак, структура интрона оказывается существенной для процессинга и транспорта РНК из ядра в цитоплазму. Механизм такого сплайсинга и его значение для дифференциальной экспрессии генов будут обсуждаться в гл. 13. Структура и функция промотораДля осуществления правильной транскрипции необходимы регуляторные элементы двух типов. Регуляторные элементы первого типа называют цис-регуляторами. Они представляют собой специфические последовательности ДНК на данной хромосоме. Цис-регуляторы оказывают действие только на ближние гены. Второй тип называют транс-регуляторами. Это растворимые молекулы (включая белки и РНК), которые продуцируются одним геном, а взаимодействуют с другими генами на той же хромосоме или на других хромосомах. Если обратиться к индукции генов в lac-опероне Е. coli, то можно вспомнить, что ген репрессора дает белок-репрессор, который взаимодействует с последовательностью оператора для генов lac-оперона. В этом случае оператор является цис-регуляторным элементом, так как он контролирует только lac-оперон своей собственной хромосомы. (Последовательность мутантного оператора на другой хромосоме может присоединять или не присоединять белок-репрессор.) Белок-репрессор, напротив, является транс-регулятором. поскольку он продуцируется одной хромосомой, а связывается с цис-регуляторным оператором на другой хромосоме (рис. 12.5). В эукариотических генах, кодирующих мРНК, обнаружены два типа цис-регуляторных последовательностей ДНК – промоторы и энхансеры («усилители»). Промоторы обычно располагаются непосредственно перед сайтом, в котором начинается |