Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ_____________________________ 145
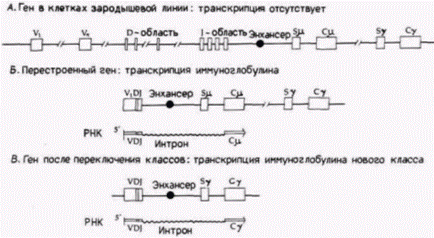
скольку для того, чтобы функционировать, промотор должен быть размещен вблизи энхансера. При переключении классов участок энхансера остается у фрагмента VDJ (рис. 12.9). Энхансеры могут объяснить также тканеспецифическую транскрипцию, так как клонированные гены иммуноглобулинов не транскрибируются после введения в ядра клеток, не являющихся В-лимфоцитами (Gillies et al., 1983; Banerji et al., 1983). Кроме того, если участок энхансера тяжелой цепи иммуноглобулина встроен в клонированный ген ß-цепи глобина, то он более чем в 100 раз стимулирует транскрипцию этого гена после его введения в клетку миеломы. Таким образом, перед нами специфический участок ДНК, который стимулирует транскрипцию лежащих поблизости генов в клетках определенного типа. Энхансеры, активность которых регулируется во времениИзвестны энхансерные последовательности, которые регулируют или время экспрессии генов, или специфичность экспрессии, или то и другое одновременно. Одно из наиболее резких по времени переключений генной активности наблюдается при переходе к средней бластуле (ПСБ) у зародышей Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.146______________ ГЛАВА 12_____________________________________________________________________________
лягушки. Было обнаружено, что определенные гены содержат энхансер, который в это время специфично активируется (Krieg, Melton, 1987). Как показано на рис. 10.34, гены, которые активируются на этой переходной стадии, можно клонировать Криг и Мелтон присоединили первые 500 пар оснований из 5’-фланкируюшей области одного из этих генов к ß-глобиновому гену Xenopus. Обычно после инъекции в оплодотворенные яйца лягушки ß-глобиновый ген не транскрибируется вплоть до поздних стадий развития. Если последовательности, лежащие с 5'-стороны от ПСБ-активируемого гена, содержат такой временной энхансер, то можно надеяться, что, начиная со времени перехода к средней бластуле, будет обнаруживаться глобиновая мРНК. Результаты экспериментов по нозерн-блот-гибридизации показали, что это действительно так. Глобиновые транскрипты отсутствовали до перехода к средней бластуле, но их можно было обнаружить после такого перехода. С помощью последовательных делений была определена локализация этого энхансера средней бластулы в последовательности длиной 74 пары оснований на расстоянии приблизительно 700 пар оснований с 5'-стороны от обычного промотора. ß-Глобиновый ген человека имеет, по-видимому, свой собственный временной энхансер, лежащий между 600-й и 900-й парами оснований после поли(А)-сайта. Если фрагмент ДНК, содержащий этот участок, помещают около γ-глобинового гена человека, то в то время, когда обычно экспрессируется ß-глобиновый ген, может активироваться ген фетального гемоглобина. (Trudel, Constantini, 1987). Тканеспецифические энхансерыПомимо энхансеров, описанных в предыдущем разделе, обнаружены также тканеспецифические энхансеры, подобные энхансеру гена тяжелой цепи иммуноглобулина. В поджелудочной железе гены экзокринных белков (химотрипсина, амилазы и трипсина) имеют энхансеры, отличающиеся от энхансеров эндокринного белка инсулина. Энхансеры обоих типов лежат в 5'-фланкирующих последовательностях соответствующих генов. Эти фланкирующие области были помещены на ген бактериальной хлорамфениколацетилтрансферазы – ген, ферментный продукт которого не обнаруживается в клетках млекопитающих (Walker et al., 1983). Затем эти гибридные гены трансфицировали в: а) клетки яичника; б) клеточную линию, секретирующую инсулин; в) линию экзокринных клеток и определяли активность маркерного фермента в клетках каждого из этих типов. Как показано на рис. 12.10. ни одна из энхансерных последовательностей не индуцировала образование фермента в клетках яичника. Однако в клетке, секретирующей инсулин, 5’-фланкирующая область инсулинового гена позволяла экспрессироваться гену хлорамфениколацетилтрансферазы, а 5’-фланкирующая об- |
 *
*