Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ_____________________________ 147
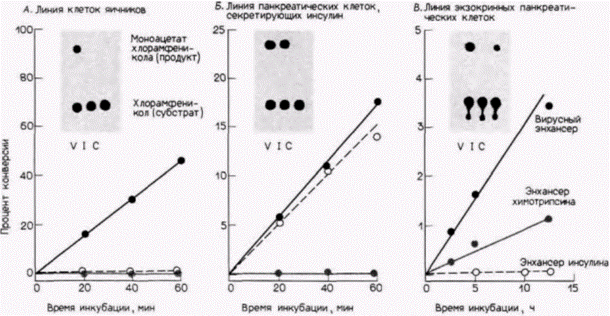
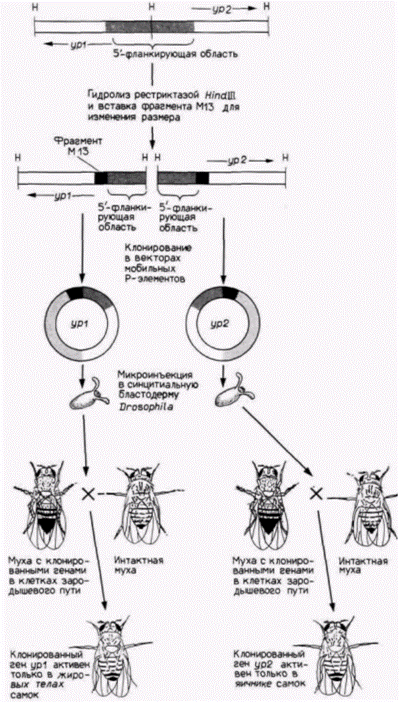
ласть гена химотрипсина не давала такой возможности. Напротив, когда клоны вводили в линию экзокринных панкреатических клеток, 5-фланкирующая последовательность для химотрипсина позволяла экспрессироваться гену хлорамфениколацетилтрансферазы, а инсулиновый энхансер не позволял. Таким образом, экспрессия генов в экзокринных и эндокринных клетках поджелудочной железы контролируется, очевидно, различными энхансерами. Нет ничего необычного в том, что один и тот же белок синтезируется в клетках не одного, а нескольких типов. Это справедливо для желточного белка, который продуцируется у зрелых самок дрозофилы. Два гена этого желточного белка – ур1 и ур2 – лежат по соседству, но транскрибируются в противоположных направлениях (рис. 12.11). Оба гена транскрибируются в яичнике и жировых телах. Эти гены могут быть отделены друг от друга с помощью рестриктазы, которая расщепляет их общую 5'-фланкирующую область (Garabedian et al., 1985). Затем каждый из этих генов может быть клонирован отдельно в векторе мобильного Р-элемента, который способен встраиваться в геном яиц дрозофилы. Прежде чем трансформировать ДНК дрозофилы этими клонами, в гены ур были введены дополнительные фрагменты плазмидной ДНК, для того чтобы продукты этих генов можно было отличить от продуктов эндогенных генов. Транскрипты с рекомбинантных клонов, подобно эндогенным мРНК, оказались специфичными в отношении пола (они присутствовали только у самок) и стадии развития (обнаруживались только у взрослых особей). Однако ур2-мРНК была обнаружена только в яичнике, а ур1-мРНК только в жировых телах. Исходные гены желточного белка должны, очевидно, иметь на своих 5'-концах два энхансера, один разрешающий транскрипцию в яичнике и другой разрешающий транскрипцию в жировых телах. Таким образом, оба гена транскрибируются в норме в клетках обоих типов. Рестриктаза, однако, разрезает ДНК между этими двумя сайтами, оставляя для yp1 энхансер, специфичный для жирового тела, а для ур2 энхансер, специфичный для яичника. Было обнаружено, что энхансер гена ур1, специфичный для жирового тела, располагается в сегменте ДНК между 196-й и 321-й парами оснований выше (с 5'-стороны от) кэп-сайта. Это определили с помощью добавления различных фрагментов 5'- Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.148_______________ ГЛАВА 12__________________________________________________________
фланкирующей последовательности к гену ß-галактозидазы E. coli. Полученные гены клонировали в Р-элементе. который затем вводили зародышам, как это было описано ранее. Потомство тех зародышей, которые включили Р-элемент, окрашивали на ß-галактозидазу (Garabedian et al., 1986). Результаты можно видеть на цветной фотографии, помещенной на одной из сторон обложки книги. В жировом теле и яичнике в отсутствие 5’-фланкирующей области ß-галактозидаза не выявляется. |
 *—
*—