Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.120_______________ ГЛАВА 11___________ ____________
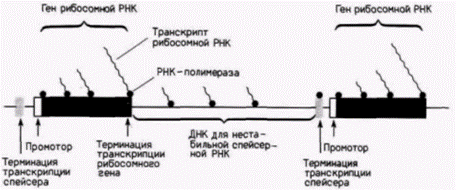
гена к другому без вхождения в пул несвязанных полимераз. Не исключено также, что свободно плавающие молекулы РНК-полимеразы I способны узнавать определенные участки ДНК в спейсерах и вовлекаться в транскрипцию рибосомной РНК. В этом случае спейсер будет действовать как «зона загрузки», накапливая новые полимеразы для транскрипции генов рибосомной РНК (Moss, 1983; Henderson, Sollner-Webb, 1986). Гены хориона дрозофилы Известно, что при синтезе белков хориона у Drosophila melanogasler гены хориона также амплифицируются. Белки хориона синтезируются фолликулярными клетками яичника. Перед началом экспрессии генов хориона весь геном фолликулярных клеток проходит дополнительные циклы синтеза ДНК, так что количество ДНК достигает 16 гаплоидных наборов. После этой репликации гены белков хориона реплицируются избирательно еще около десяти раз. Эта амплификация происходит только в фолликулярных клетках яичника (Spradling, Mahowald, 1980).
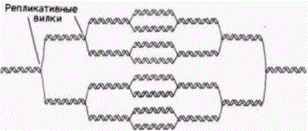
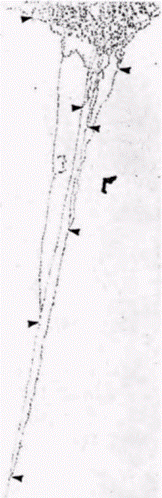
Амплификация обусловлена дополнительными циклами репликации ДНК лишь в определенных областях генома (Spradling, 1981). Эти области ДНК характеризуются наличием нескольких ветвей, каждая из которых содержит амплифицированные гены хориона (рис. 11.14 и 11.15). В данном случае амплификация происходит, но гены остаются присоединенными к хромосомам. Селективная транскрипция геновБольшинство генов не используют избирательную амплификацию в качестве механизма для обеспечения крупномасштабной экспрессии. В развивающихся эритроцитах, в которых гемоглобин составляет 98% всего синтезируемого белка, не наблюдается избирательной амплификации глобиновых генов. В шелкоотделительной железе гусениц, продуцирующих огромное количество белка шелка фиброина, весь геном амплифицируется с помощью политении. Даже в этом случае отсутствует селективная амплификация генов фиброина. На основании этого мы полагаем, что определенные гены могут транскрибироваться избирательно. Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ ИЗМЕНЕНИЕ ТРАНСКРИПЦИИ В ХОДЕ РАЗВИТИЯ_______________________________________ 121
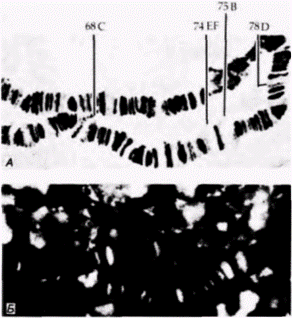
Хромосомные пуфы и хромосомы типа ламповых щетокПрирода политенных хромосом и их пуфов была рассмотрена в гл. 10. Представленные в ней данные иллюстрируют контроль за активностью определенных генов на уровне транскрипции. Если к слюнным железам добавляют экдистерон, то наблюдается возникновение одних пуфов и исчезновение других. Образование пуфов опосредовано присоединением экдистерона к специфическим участкам на хромосомах. Это может быть продемонстрировано путем «сшивания» экдистерона с хроматином, которое фиксирует его положение на местах связывания. Затем добавляют кроличьи антитела против экдистерона и несвязавшиеся антитела удаляют промывкой. В конце добавляют меченные флуоресцеином козьи антитела, полученные против иммуноглобулинов кролика, и несвязывавшиеся антитела отмывают. В результате флуоресцентная метка должна обнаруживаться в тех местах, где козьи антитела связались кроличьими антителами, а кроличьи антитела должны присоединяться только к экдистерону. Таким образом, флуоресцентная метка должна появиться на хромосоме в каждом сайте, где был связан экдистерон (Gronemeyer, Pongs, 1980). Результаты свидетельствуют о том (рис. 11.16), что практически все экдистерон-чувствительные пуфы связывают экдистерон. Экдистерон-чувствительные пуфы, обнаруживаемые у поздней личинки третьего возраста (когда она готовится стать куколкой), можно грубо разделить на три категории. Пуфы, которые исчезают в присутствии экдистерона; пуфы, которые быстро индуцируются экдистероном, и пуфы, которые появляются через несколько часов после стимуляции экдистероном. Ранние пуфы формируются в пределах 1 ч после добавления экдистерона. Кроме |