Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ___________________ 207
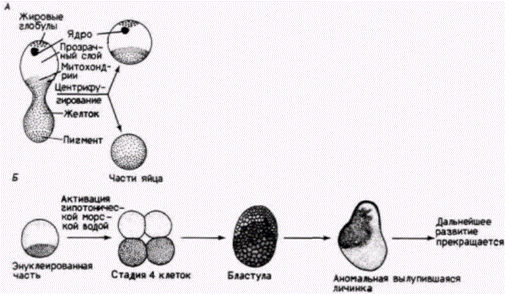
Вторая группа данных, свидетельствующих о материнской регуляции раннего развития, получена в работах по энуклеации. Если после оплодотворения яйца из него удалить ядро, то можно выяснить, как далеко проходит развитие в отсутствие синтеза новых мРНК. Энуклеация проводилась с помощью физических, химических и генетических методов. Физическую энуклеацию ооцита осуществила Харвей (Harvey, 1940). Она помещала неоплодотворенные яйцеклетки морского ежа в раствор сахарозы, имеющий ту же плотность, что и сами яйцеклетки. При центрифугировании суспензия расслаивалась, и в конечном счете яйцеклетки разрывались на две глобулы (рис. 14 11. А). Легкая часть содержала ядро, а тяжелая часть оказывалась энуклеированной. Затем Харвей партеногенетически активировала энуклеированные части, поместив их в гипотоническую морскую воду. Эти части («партеногенетические мерогоны») дробились, образовывали аномальную бластулу (лишенную бластоцеля) и благополучно выклевывались (рис. 14.11,5). Дальше развитие прекращалось. Результаты аналогичных экспериментов с использованием ооцитов лягушки также показали, что дробление может происходить даже при полном отсутствии хромосом (в том случае, если в икру были инъецированы центриоли для компенсации тех, которые в норме поставляются спермием). В этом случае также возникает аномальная бластула. Опыты по физической энуклеации показывают, что определенные яйцеклетки даже в отсутствие ядра способны проходить в своем развитии стадию средней бластулы. Запасенные мРНКДанные о том, что ооцит контролирует раннее развитие благодаря запасенной им мРНК, были впервые получены двумя группами исследователей (Brachet et al., 1963; Denny, Tyler, 1964). Результаты этих исследований показали, что энуклеированные и активированные части яйцеклеток морского ежа содержат РНК и могут синтезировать белки со скоростью, сравнимой со скоростью синтеза в нормально оплодотворенных яйцах. В энуклеированных ооцитах транскрипции происходить не может, следовательно, для синтеза белков в них должна использоваться запасенная мРНК. Было показано, что прирост в белковом синтезе не может быть обусловлен транскрипцией митохондриальной ДНК ооцита (Craig, Piatigorsky, 1971). Представляется, что ооцит создал запас мРНК, которые не транслируются до оплодотворения. Мягкие способы энуклеации стали возможны, когда было открыто, что антибиотик актиномицин D ингибирует синтез РНК, и транскрипция может быть выключена путем помещения свежеоплодотворенной яйцеклетки в раствор этого антибиотика. Когда ооциты морского ежа обрабатывали актиномицином D в концентрации, достаточной для Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.208_______________ ГЛАВА 14________________________________________________________________________
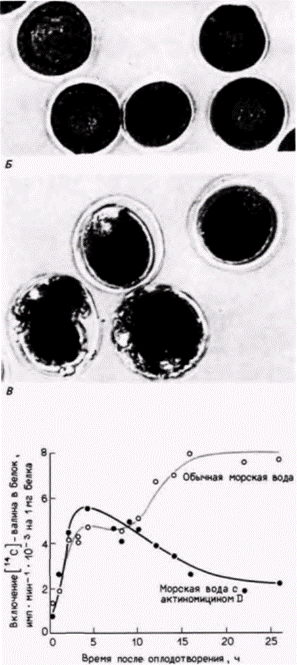
подавления синтеза РНК в ней на 94%, зародыши в своем развитии все еще достигали стадии бластулы (рис. 14.12. Б) (Gross, Cousineau, 1964). То, что это развитие зависит от запасенных мРНК, а не от предварительно синтезированных белков, было показано с помощью обработки оплодотворенных ооцитов эметином или циклогексимидом. Эти антибиотики ингибируют трансляцию, и зародыши, оплодотворенные в присутствии этих ингибиторов, не могут развиваться вовсе (рис. 14.12. В). В ряде исследований был продемонстрирован всплеск белкового синтеза вскоре после оплодотворения. Результаты экспериментов Гросса и его коллег с использованием актиномицина D показали, что в химически энуклеированных зародышах синтез белка после оплодотворения увеличивается точно так же, как в контрольных (рис. 14.13). Однако спустя 6-10 ч в обработанных зародышах наблюдается спад белкового синтеза и второй всплеск белкового синтеза на стадии бластулы у них не происходит. Молекулы мРНК для этого второго подъема синтеза белка транскрибируются в ядре. Таким образом, зародыши, обработанные актиномицином D, могут развиваться вплоть до выклева на стадии бластулы, а далее развитие останавливается. Следовательно, запасенных мРНК ооцита
|